Article Text

Abstract
Glycosylation is a key modification of proteins and lipids and is involved in most intermolecular and intercellular interactions. The gastrointestinal mucus gel is continuous and can be divided into two layers: a secreted loosely associated layer and a layer firmly attached to the mucosa. In addition, the membrane-bound glycosylated proteins and lipids create a glycocalyx, which remains adherent on each cell and is dynamic and responsive to the physiological state and environment of the cell. The secreted glycans form a mucus gel layer that serves as a physicochemical sensor and barrier network and is primarily composed of mucins and associated peptides. These glycans protect gut epithelial cells from chemical, biological and physical insults and are continuously renewed. Pathogens colonise and invade the host epithelial cells using protein–protein and glycan–lectin interactions. During the process of colonisation and infection, the glycosylation state of both host and pathogen change in response to the presence of the other. This complex modulation of glycan expression critically determines pathogenesis and the host response in terms of structural changes and immune response. In addition, by influencing host immunity and gut glycosylation, the microbiota can further effect protection against pathogens. In this review, the roles of host glycosylation in interactions with two prevalent bacterial pathogens, Campylobater jejuni and Helicobacter pylori, are discussed to illustrate important concepts in pathogenesis.
- Glycosylation
- mucins
- bacterial pathogenesis
- Campylobacter jejuni
- Helicobacter pylori
- bacterial adherence
- bacterial pathogenesis
- glycobiology
- sugars
Statistics from Altmetric.com
- Glycosylation
- mucins
- bacterial pathogenesis
- Campylobacter jejuni
- Helicobacter pylori
- bacterial adherence
- bacterial pathogenesis
- glycobiology
- sugars
Introduction
Beyond genes and proteins
For the past 60 years, the central dogma of molecular biology has emphasised the importance of the transcription of DNA to RNA and translation of RNA into amino acids to form proteins. By their physical and chemical nature, DNA, RNA and proteins are linear molecules and are synthesised using a pre-existing template. However, with the completion of sequencing of several organisms' genomes, it has emerged that the complement of genes alone does not define the complexity of an organism and its interactions with other organisms. Once amino acid chains are synthesised, they can undergo over 200 covalent modifications, so-called post-translational modifications, which determine the biological, physical and chemical properties of the proteins.1 The realisation that proteins are heavily modified, and that these modifications are critical to their structure and function, has led to a shift in emphasis from primarily studying genes and proteins to the analysis of post-translational modifications, such as phosphorylation, sulfation and glycosylation.
What is glycosylation?
Glycosylation is one of the most prevalent modifications mediated by complex enzymatic machinery, whereby glycans (sugars) are covalently attached to specific amino acid sites of proteins. Mammalian glycosylation takes place on nascent protein chains when they are passing through the endoplasmic reticulum (ER) and the Golgi complex. Subsequently, these proteins and lipids are transported to different organelles in the cell, to the cell membrane, or secreted from the cell.
All cells, from bacteria to human, are covered in glycans. Glycosylation leads to the formation of monomeric and multimeric glycan linkages which are essential for cell viability, biochemical communication and normal function. The resulting glycome encompasses a diverse and abundant repertoire of glycans, which form one of the four fundamental macromolecular components of all cells (together with nucleic acids, proteins and lipids). Glycans have important biological functions in protein maturation and turnover, cell adhesion and trafficking, and receptor binding and activation. Protein glycosylation has crucial roles in most physiological processes and diseases, including cell signalling, cellular differentiation and adhesion. Dysfunction leads to pathogenesis of infectious diseases and development of chronic diseases, for example, cancer, neurological disorders and severe inflammation.2
The diversity and complex characteristic of glycans on glycoconjugates is derived from the many ways in which monosaccharides can be linked together during their synthesis to form higher-order structures (figure 1B). Unlike nucleic acids and proteins, which are synthesised on the basis of a template and are linear molecules, glycan synthesis is not template-based and glycans can be both linear and branched. They can produce a large number of isomers because of the possibility of linkage and anomeric isomers. These isomers have very different three-dimensional structures and biological activities. A monosaccharide can be involved in more than two glycosidic linkages, thus serving as a branch point. The common occurrence of branched sequences is unique to glycans and contributes to their structural diversity. The glycosidic linkage is the most flexible part of a disaccharide structure. Thus, a disaccharide of well-defined primary structure can adopt multiple conformations in solution that differ in the relative orientation of the two monosaccharides. The combination of structural rigidity and flexibility is typical of complex carbohydrates and, more than likely, essential to their biological function.2 3

Glycosylation on the mammalian cell surface. (A) Diagrammatic representation of glycosylation on glycoconjugates of the mammalian cell surface including glycolipid, proteoglycan, N-linked and O-linked glycoprotein molecules. (B) The types of mammalian glycans, their diversity and complexity. Glycan structures of the six classes of secretory glycan (N-glycans, hyaluronan, O-glycans, glycolipids, glycosylphosphatidylinositol (GPI) anchor and glycosaminoglycans) and the single intracellular glycan, O-linked β-N-acetylglucosamine (O-GlcNAc), are shown. Representative examples of each type are indicated using the symbol nomenclature for monosaccharides (see symbols). PI is phosphatidylinositol, Et–P denotes a phosphoethanolamine linkage, and –s denotes the sulfation positions of the glycosaminoglycan chains.
Glycans are linked to other biomolecules, such as lipids or amino acids within polypeptides, through glycosidic linkages to form glycoconjugates. The glycan may be a single monosaccharide or an oligosaccharide. The attachment of many glycans to a polypeptide scaffold creates tremendous diversity among glycoproteins (figure 1). Glycoconjugates therefore make up a highly dynamic system capable of participating in a wide range of biological interactions.3
Nature of sugars and glycosylation—glycoproteins, proteoglycans and glycolipids
Mammalian glycoconjugates are classified on the basis of whether the glycans are attached to proteins or lipids (figure 1A). In the case of protein glycosylation, there are two major types of glycoprotein, N-linked and O-linked (figure 1B). In N-linked glycoproteins, the glycosylation takes place on an asparagine residue when asparagine is found in the following consensus motif: Asn-X-Ser/Thr (where X can be any amino acid but proline). N-Linked glycoproteins go through a complex series of attachment events in the ER and the Golgi complex, with the resultant glycoprotein carrying a mixture of glycan structures varying in length from five to 15 sugars. These structures are primarily composed of N-acetyl-d-glucosamine (GlcNAc), d-mannose (Man), d-galactose (Gal), l-fucose (Fuc) and sialic acid also known as neuraminic acid (N-acetylneuraminic acid; Neu5Ac) (figure 1B). Several cell-surface-bound as well as secreted proteins are N-glycosylated—for example, erythropoietin and immunoglobulins.3 Unlike N-linked glycosylation, O-linked glycosylation takes place entirely in the Golgi complex where a single N-acetyl-d-galactosamine (GalNAc) residue is enzymatically attached to the Ser/Thr residue on folded proteins (figure 1B). In general, O-linked glycan chains are shorter than N-linked glycan chains but are composed of the same monosaccharides as used in N-linked glycosylation. Cell-surface-bound and secreted mucins are examples of O-linked glycoproteins (figure 2).4 These heavily O-glycosylated proteins carry clusters of GalNAc-based glycans in repetitive Ser- and Thr-rich VNTR (variable number of tandem repeats) domains. These complex oligosaccharide structures have been arbitrarily assigned three main domains: the core region, which comprises the innermost two or three sugars of the glycan chain proximal to the peptide; the backbone region, which mainly contributes to the length of the chains formed by uniform elongation; and the peripheral or terminal region, which exhibits a high degree of structural complexity (figure 2).4 Mucins are the most prominent host molecules involved in host–pathogen interactions and are discussed in more detail in the following sections.

Mucin structure and glycosylation. (A) Schematic representation of structures of secreted and cell-membrane-associated mucins showing different domains. (B) Cartoon showing distribution and structure of gel-forming secreted, non-gel-forming secreted, and membrane-bound mucins. (C) Schematic representation of the complex glycosylated structure and multimeric assembly of a gel-forming secreted mucin. Immediately after its synthesis and translocation into the endoplasmic reticulum, the mucin polypeptide chain is N-glycosylated and forms disulfide-bonded dimers through its C-terminal Cys-rich domains. The dimers are then transported to the Golgi complex, and the tandem repeat domains are O-glycosylated. Once the glycosylated dimers reach the trans-Golgi compartments, they are assembled into disulfide-bonded multimers through their N-terminal D-domains. The Y-shaped structures on the N-terminal D-domains and the C-terminal Cys-rich domains represent N-linked oligosaccharides. The squiggly lines perpendicular to the polypeptide chain represent O-linked oligosaccharides in the tandem repeat domains.
Proteoglycans are a subclass of glycoproteins, but the distinction between the two is not very clear, as both carry a protein backbone with glycans attached at specific amino acids. In the case of proteoglycans, several acidic disaccharide-repeating units are attached to the protein backbone. These disaccharide units (called glycosaminoglycans) are composed of a hexosamine and a hexose or a hexuronic acid (glucouronic acid or iduronic acid). Proteoglycans are integral components of the extracellular matrix; examples include chondroitin sulfate, dermatan sulfate and heparan sulfate.5
A glycolipid (also referred to as glycosphingolipid) is a glycoconjugate in which glycan chains are attached to the lipid ceramide with glucose (Glc) or Gal as the first sugar, thereby forming glucosylceramide and galactosylceramide. Glycolipids are ubiquitous components of membranes, and their distribution and structures vary depending on the location in the organism.6
During the process of colonisation and infection, the glycosylation state of both host and pathogen change in response to the presence of the other. This complex modulation of glycan expression critically determines pathogenesis and the host response in terms of structural changes and immune response. The aim of this paper is to review the roles of glycosylation of the gastrointestinal (GI) tract in colonisation and invasion of this mucosa by bacteria. In particular, the gastroduodenal pathogen, Helicobacter pylori, and the enteric pathogen, Campylobacter jejuni, will be used as examples of acute and chronic bacterial infections, respectively, to illustrate important concepts concerning the role of GI glycosylation in pathogenesis.
Mucus layer and epithelial glycocalyx as barriers to bacterial infection
Mucus layer
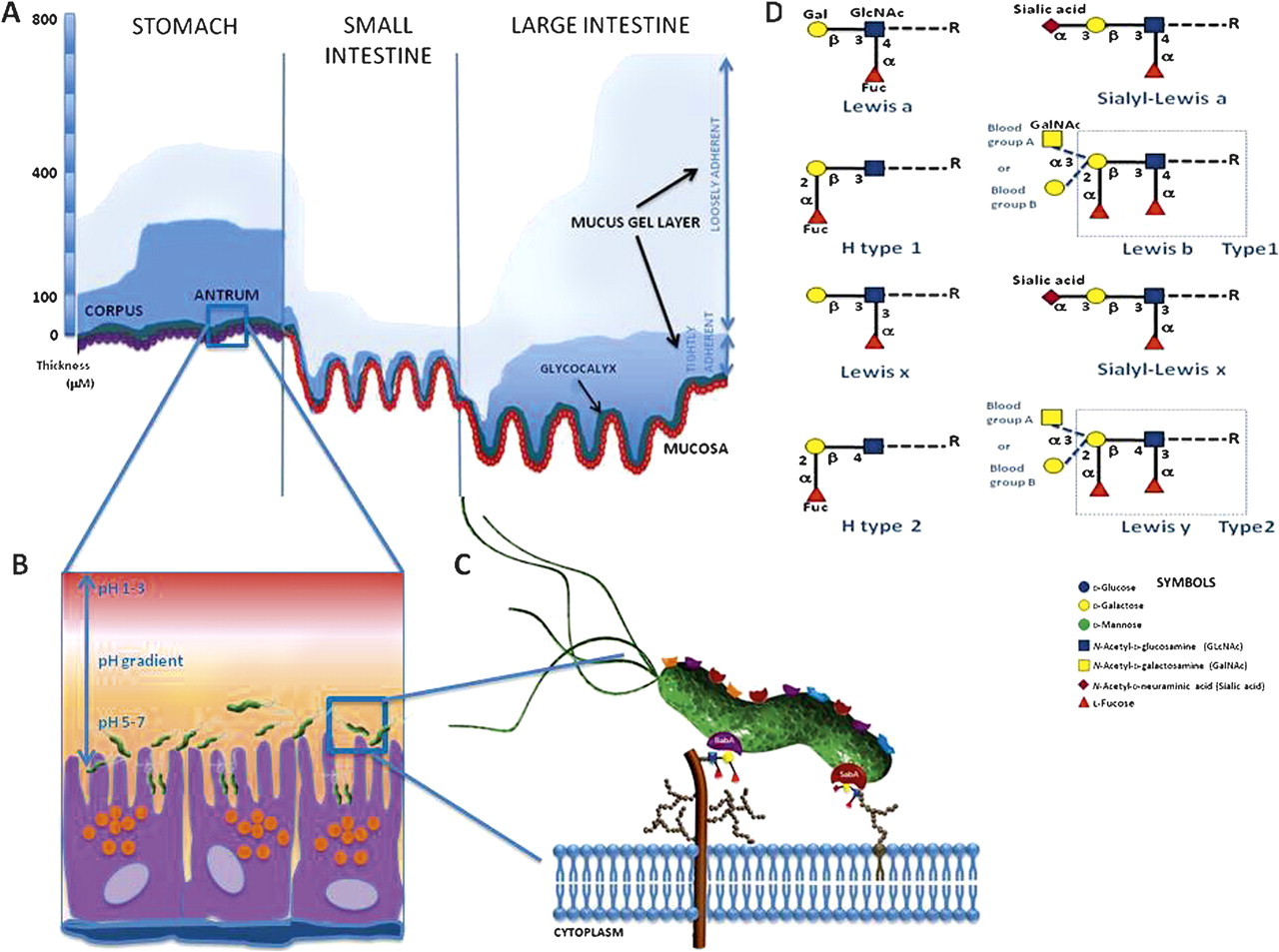
The mucosal surface of the GI tract, which is a complex ecosystem of resident microbiota, immune cells and an epithelium covered by a mucus layer, is the largest body surface (200–300 m2) in contact with the external environment.7 The adherent gastrointestinal mucus gel in vivo is continuous and can be divided into two layers: a loosely adherent layer and a layer firmly attached to the mucosa (figure 3A). It ranges in thickness from 300 μm in the stomach and 700 μm in the intestine,8 and represents the front-line defence barrier between the external environment and tissues of the host.9 The relative thickness of the two component layers of the mucus gel varies for different regions of the gut. Thus, in the stomach, the thickness of the firmly adherent mucus component is 80 and 154 μm for the corpus and antrum, respectively, with an overlying layer of loosely adherent mucus of similar thickness (figure 3A). The gastric pathogen H pylori has been shown to reduce the rate of renewal of the mucus layer, as well as cause an acid-induced increase in its renewal rate. The mucus gel is the thickest in the colon, and the rate of mucus accumulation is highest. In the small intestine, the mucus layer is thin and discontinuous.8 9 This viscous, supramucosal gel presents a highly hydrated, mechanical and lubricative barrier, as well as a matrix for immune-related and antimicrobial proteins. It is not static, since the outermost loosely adherent mucus layer is continually removed by movement of the luminal contents of the GI tract and renewed. The rate of renewal of the mucus layer varies in different parts of the GI tract (figure 3A).8 9 Thus, the mucus layer protects the underlying epithelial cells against chemical, enzymatic, microbial and mechanical insult, and serves as a physical gel to inhibit and entrap invading microbes and aid clearance. In addition, in the stomach, mucus acts as an unstirred layer in which bicarbonate ions secreted by the surface epithelium counteract protons diffusing from the lumen into the gel, causing the pH in the mucus layer to range from acidic in the lumen to neutral at the cell surface which, in turn, can influence bacterial colonisation (figure 3B).10 For example, H pylori usually resides within the one-fourth zone (0–25 μm) of mucin gel above the epithelium in close proximity to the cells where the pH ranges from 5 to 7 (figure 3B).10
{kind=link}
{kind=link}
{kind=link}
The gastrointestinal mucus layer and its role in bacterial adhesion. (A) Diagram showing the thickness of the two mucus gel layers in vivo in the stomach and small and large intestine. The mucus gel layer is continuous throughout the gastrointestinal tract and does not follow the contours of the villi in the intestine. The membrane-associated glycocalyx is also shown. (B,C) Glycan-mediated adhesion of Helicobacter pylori to gastric epithelial cells. The figure illustrates some of the characterised bacterial–glycan interactions that contribute to successful colonisation of the gastric mucosa. (D) Schematic representation of the terminal ABH structures and Lewis epitopes present in O-glycans, which act as receptors for H pylori adhesins, BabA and SabA.
The protective physicochemical properties of mucus are substantially attributable to the high carbohydrate content of mucins, which also presents considerable potential for interaction between microbial lectins and O-linked glycans. In addition, glycosidases and other enzyme activities secreted or induced by microbes have the capacity to directly modify these glycans after the mucins have been secreted or exported to the cell membrane.11 12 This strategy is significant as a mechanism for generating binding ligands and in the provision of sugars for bacterial metabolism.13 Bacterial interaction with the supramucosal gel can lead to chronic colonisation of the mucus. On one hand, the mucosal microbiota blocks access to the underlying cells, and on the other, in the case of pathogens, necessitates the engagement of gel-penetration strategies before cellular adhesion.9 It is therefore surprising that the relevance of this layer to colonisation and invasion of the mucosal epithelium has not historically been of higher priority.
Glycocalyx
Underneath the mucus layer, the second line of defence is constituted by the epithelial glycocalyx, which is partially integrated with the overlying gel (figure 3A).14 The glycocalyx represents an array of highly diverse glycoproteins and glycolipids expressed on the membrane of epithelial cells. The glycocalyx is highly variable from tissue to tissue as well as in thickness (intestinal microvilli tips, 100–500 nm; lateral microvilli surface, 30–60 nm) (figure 3A).8 The glycosylation machinery of mucosal epithelia not only varies with the cell type but is also influenced by the sub- and supra-mucosal environment—for example, hormonal status, inflammation and microbial colonisation have all been shown to influence the glycosylation of epithelial glycoproteins.15–18 Like the mucus gel, the glycocalyx is constantly renewed; the average turnover time of the human jejunal glycocalyx is 6–12 h. In the gut lumen, the glycocalyx interacts with the overlying mucus layer, the gastric-biliary juices and the resident probiotic microflora to limit colonisation by pathogens. The complex glycosylated structures expressed within the glycocalyx serve as receptors and form the primary site for bacterial adhesion. For example, the carcinoembryonic antigen-related cell adhesion molecule 6 (CEACAM6) expressed on the apical surface of ileal epithelial cells serves as a receptor for adherent-invasive Escherichia coli in Crohn disease.19 Expression of such glycosylated ligands can in turn be modulated by bacterial infection and inflammation, further promoting colonisation.
Bacterial colonisation and adherence
To colonise mucosal surfaces and invade the epithelium, microbes commonly interact with glycan structures of the host glycocalyx.11 16 Bacterial fimbriae (or pili), various outer membrane proteins and cell wall components—for example, lipopolysaccharides (LPSs)—may all function as adhesins. Adhesion to host cells is considered to be mediated by hydrophobic interactions, cation-bridging whereby divalent cations counteract the repulsion of the negatively charged surfaces of bacteria and host, and receptor-ligand binding.7 11 One of the most extensively studied mechanisms of adhesion is the binding of bacterial lectins to the corresponding glycosylated receptors associated with host cells.10 Bacteria may have multiple adhesins with different carbohydrate specificities, and modulation of surface receptor density, kinetic parameters, or topographical distributions of these receptors on cell membranes regulate adhesion. After H pylori enters the stomach, infection occurs in three crucial steps: establishing colonisation; evading host immunity; and invading gastric mucosa. Urease activity and motility mediated by the flagella of H pylori are important in penetrating the mucus gel and harbouring colonies beneath the gastric mucus in niches adjacent to the epithelium. Several putative adhesins attach the organism to the gastric epithelium and prompt the succeeding processes for evading host immunity and invading the mucosa.20 21 H pylori expresses at least six lectin-like adhesins for recognition of the gastric mucosa, of which the blood group-binding adhesin (BabA), which binds to Lewis b (Leb) and H-1 antigens,20 and sialic acid-binding adhesin (SabA), which binds sialyl-Lex in inflamed tissue,21 are most important for colonisation (figure 3C). Human Lewis antigens represent terminal modifications on mucin-type O-glycans. Their structure is composed of two different backbones (figure 3D). The type 1 chain contains Gal and GlcNAc composed as Gal-β(1,3)-GlcNAc, while the type 2 chain has Gal-β(1,4)-GlcNAc, which is also termed N-acetyl-lactosamine (LacNAc). Adding α-Fuc as a side chain to the backbone structure on the 1–4 location, on both 1–2 and 1–4 locations, and on both 2–2 and 2–4 locations gives rise to Lewis a (Lea), Leb and sialyl-Lea antigens, respectively (figure 3D). Based on the backbone structure of the type 2 chain, the addition of α-Fuc as a side chain at the 1–3 location on both 1–2 and 1–3 locations, or on both the 2–2 and 2–3 locations, gives rise to the Lewis x (Lex), Lewis y (Ley) and sialyl-Lex antigens, respectively (figure 3D).22
The distribution of type 1 antigens such as Lea and Leb is primarily on the surface of the gastric epithelium. This is the primary site of ligation for the H pylori adhesion, BabA. BabA-positive status and its binding to Leb have been associated with severe gastric injury, high H pylori density, and severe clinical outcomes.23 The fucosylated H-type 1 and Leb antigens are naturally expressed on the gastric mucosa of secretor or Lewis-positive individuals. The secretor status depends on an active FUT2 enzyme, an α(1,2)-fucosyltransferase that catalyses the addition of terminal α(1,2)-fucose residues. Inactivating mutations in the secretor gene affect 20% of the human population (known as non-secretors) and have been associated with reduced susceptibility to infections by H pylori.23 24 Nearly 95% of the BabA-encoding H pylori adapt to any of the Leb sub-types, and such strains are termed ‘generalists’. A small subset of H pylori strains express BabA, but cannot adhere to the A-Leb antigen. Such strains are called ‘specialist’.23 BabA can also act as a mediator for H pylori binding to MUC5AC, even in non-secretors or those without Leb. In contrast, type 2 antigens may be located deeper in the glands, such as mucous, chief, or parietal cells. Leb is weakly expressed on the surface epithelium during chronic inflammation, as well as in gastric atrophy (atrophic gastritis) and intestinal metaplasia. Under these clinical conditions, there is an upward migration of Lex associated with atrophy of the epithelium. During chronic inflammation or in atrophic gastritis, H pylori binds to Lex and sialyl-Lex via SabA adhesin. SabA production and SabA-mediated adhesion are associated with severe disease outcomes, including gastric atrophy, intestinal metaplasia and the development of gastric cancer. The minimal structure required for SabA adhesin binding was shown to be NeuAcα2-3Gal (figure 3C,D).25
H pylori CagL binds to the integrin, α5β1, to form an injection pile by which CagA translocates into the cytosol of gastric epithelium, using a type IV secretory system. Other known H pylori adhesins include neutrophil-activating protein, which binds to sulfated carbohydrate structures,26 27 a 25 kDa adhesin that binds the glycoprotein laminin in the extracellular matrix,28 29 as well as adherence-associated lipoprotein A and B (AlpA/B)10 30 and HorB,31 whose gastric receptors have not been identified. Moreover, H pylori lectins bind molecules in the oral cavity, such as salivary agglutinin, that may influence re-infection of the gastric mucosa from this secondary infection reservoir.32 At least four protein adhesins, which differ substantially in activity and importance depending on the anatomical site within the orogastric route, the mucin-type, the environmental pH and the gastric disease status, are involved in H pylori binding to mucus-related oligosaccharides.33 The adhesive interactions are not limited to protein adhesins, as H pylori LPS interacts and binds with the gastric epithelium34 through recognition by the host lectin, galectin-3.35 As occurs with protein adhesins,36 environmental pH, particularly low pH, can influence expression of the LPS ligand involved in the latter interaction.37 In addition, this bacterium recognises a range of glycosylated molecules, not only glycoproteins and oligosaccharide moieties, but also proteoglycans and glycolipids.10 In contrast, the nature and specificities of the lectins expressed by intestinal-colonising C jejuni appear less diverse,38 39 possibly reflecting the lesser mucus and tissue diversity of the jejunal environment (figure 3A). Nevertheless, although individual adhesion–receptor binding is usually of low affinity, clustering of adhesins and receptors can cause multivalency effects resulting in strong attachment.
The O antigen of H pylori LPS shares structural homology with the Lewis blood group antigens expressed in human gastric mucosa, including both type 1 chains, namely Lea and H-type 1, and type 2 chains, such as Lex and Ley. The expression of these antigens undergoes phase variation, with different bacterial fucosyltransferases being switched on and off, contributing to a dynamic glycosylation even in the same host. It has been proposed that Lex structures expressed by the H pylori O antigen side chain may further promote adhesion to gastric epithelial cells.37 40
Collectively, ligand–receptor binding can have different consequences for bacterial pathogens during colonisation of the GI tract and whether the resultant infection is acute or chronic. Adhesion can affect bacteria by stimulation/inhibition of growth, as well as induction of other adhesive structures and proteins required for invasion, whereas the effects of adhesion on host cells can be altered morphology, fluid loss, induction of cytokine release, upregulation of cell adhesion molecules, and apoptosis.7 In addition, infection and associated inflammation can induce changes in mucosal glycosylation,41 42 including the release of carbohydrates for bacterial utilisation. An example of this occurs in H pylori infection whereby human α-l-fucosidase is secreted which liberates l-Fuc from host cells, which becomes available for H pylori to synthesise mammalian Lewis antigens,43 which contribute to pathogenesis of the bacterium.40 44
Of the range of GI barrier-related molecules with which bacterial pathogens interact, mucins as a common family of molecules present in secreted mucus and in the glycocalyx are receiving increasing attention and will form the predominant focus for further discussion in this review.
Properties and characteristics of mucins
General attributes of mucins
The polymeric scaffolding of mucus is formed by secreted mucin glycoproteins. However, it is important to note that certain mucins (eg, MUC1) can be membrane-bound and hence form a major constituent of the epithelial glycocalyx of a variety of mucosal tissues (figure 2B). Mucin-type molecules consist of a core protein moiety (apomucin) within which a number of carbohydrate chains are attached to Ser, Thr and Pro residues by glycoside bonds forming O- and N-linked oligosaccharides15 45 (figure 2C). Mucins have a high molecular mass (5×105 to 30 ×106), and more than 50% (often 70–80%) of the mucin molecular mass is composed of carbohydrate.9 Each mucin carries of the order of 100 different oligosaccharide structures. These carbohydrate chains are often clustered into highly glycosylated domains, giving the mucin a ‘bottle-brush’ appearance (figure 2C). In secreted mucins, these O-linked oligosaccharides are concentrated in large peptide domains of repeating amino acid sequences rich in Ser and Pro.15 The size and number of repeats vary between mucin types, and in many of the mucin genes there is polymorphism in the number of repeats (VNTR) (table 1)8 9 (figure 2C). Thus, the size of such mucins can differ substantially between individuals and could represent a basis for differences in infection susceptibility between hosts.
Distribution of mucins in the gastrointestinal tract and in associated tissues
Mucin categories
Mucins can be divided into three distinct subfamilies: (i) secreted gel-forming mucins; (ii) cell surface mucins; (iii) secreted non-gel-forming mucins (table 1). The gel-forming mucins (eg, MUC2, MUC5AC, MUC6 and MUC19) are a major constituent of mucus and confer its viscoelastic properties. Gel-forming mucins contain N- and C-terminal Cys-rich domains, which are involved in homo-oligomerisation mediated by intermolecular disulfide bonds, which is likely to result in web-like supramolecular structures critical for the rheological properties of the mucus gel4 8 46 47 (figure 2C). The extensive O-glycosylation of secreted mucins gives protection from proteolytic enzymes and, because of the dense glycosylation, induces a relatively extended conformation, which causes these secreted oligomeric molecules to occupy large volumes equivalent in size to small bacteria.48 In addition to the secreted mucins, cell surface mucin glycoproteins (eg, MUC1, MUC3A/B, MUC4, MUC12 and MUC13) are expressed on the apical membrane of all mucosal epithelial cells, representing major constituents of the glycocalyx8 15 (figure 2A). They are monomeric and may be shed into the overlying mucus and contribute to viscosity but not elasticity of the gel.9 In general, their large VNTR domains (table 1) predict that they form rigid elongated structures, which together with their high expression, indicate they are a predominant constituent of the glycocalyx. Cell surface mucins associate with the cell membrane by an integral transmembrane domain and possess short cytoplasmic tails that associate with cell cytoskeletal proteins and are important for transducing external signalling into the mammalian cell (eg, with MUC1 and MUC4).7 17 The secreted non-oligomerising mucins include the MUC7 salivary mucin (table 1).
Mucin glycosylation
The oligosaccharide content and carbohydrate structures of mucins vary according to cell lineage, tissue location and developmental stage. O-Linked glycans contain 1–20 residues, which occur both as linear and branched structures (table 2). The carbohydrate chain is initiated with a GalNAc residue linked to Ser or Thr, to which is added the core structures followed by the backbone region produced by elongation units of either type 1 or type 2 LacNAc.9 15 The chains are terminated by Fuc, Gal, GalNAc or Neu5Ac residues in the peripheral region, forming histo-blood-group antigens such as A, B, H, Lea, Leb, Lex, Ley, as well as sialyl-Lea and sialyl-Lex structures (figure 3D).22 44 The carbohydrate structures present depend on which glycosyltransferases are expressed in the cells—that is, by the genotype of the individual.22 The H-1 structure is made by the secretor (Se) gene product; the majority of humans worldwide carry this structure and are thus referred to as secretors. Individuals may also express the Lewis gene (90% of the Caucasian population), and, provided that they are also secretors, will modify the H-1 antigen to Leb, whereas if they are non-secretors, Lea will be expressed on type 1 chains. Also, mucins contain a smaller number of N-linked oligosaccharides that have been implicated in folding, oligomerisation or surface localisation8 (figure 2C).
Common O-linked oligosaccharide structures on mucins
The terminal structures of mucin oligosaccharides are highly heterogeneous and vary between/within species and even with tissue location within a single individual. For example, in the gastric mucosa, Lea and Leb blood-group antigens mainly appear on the surface epithelium on MUC5AC, whereas the Lex and Ley antigens are expressed in mucous, chief and parietal cells of the glands, co-localised with MUC6.49 50 Also, mucin glycosylation changes with infection progress; mucins from a healthy human stomach express low levels of sialic acid and sulfate and are therefore predominantly neutral, but prolonged infection with H pylori decreases fucosylation and increases sialylation of mucins.21 51 However, the sialylation pattern returns to normal after successful H pylori eradication therapy. The structural diversity of glycosylation has been suggested to allow the host to cope with diverse and changing pathogen challenges.11 Notably, host susceptibility to a specific bacterial pathogen and subsequent disease severity can vary between individuals expressing different histo-blood groups. This has been demonstrated for H pylori infection in rhesus monkeys, which shares strong similarities to the natural history of H pylori infection in humans.24 This bacterium has an ability to vary and adapt its surface characteristics, including expression of glycosylation and outer membrane proteins to aid infection in humans and in animal models.35 52–55 A strong correlation has been found between the adhesive properties of H pylori endemic in specific human populations and the mucin-related blood group structures expressed by those populations,51 as well as the ability of the bacterial adhesins to vary to aid colonisation.10 23 56
Changes in the expression levels and glycosylation patterns of mucins have been associated with several diseases, including carcinomas. In gastric cancer, alterations in mucin polypeptide expression have been reported: loss of expression of MUC5AC, increased mucin heterogeneity, and glycosylation changes, including exposure of simple mucin-type carbohydrates.57 The majority of gastric carcinomas are preceded by a precancerous stage, characterised by sequential development of atrophic gastritis leading to intestinal metaplasia, and dysplasia. Intestinal metaplasia consists of replacement of the gastric mucosa by an epithelium that histologically resembles the intestinal mucosa and is characterised by the presence of absorptive cells, Paneth cells, and goblet cells secreting sialo-mucins and/or sulfo-mucins and corresponds to the small intestine phenotype.58 Altered mucin expression patterns reported in intestinal metaplasia include underexpression of MUC1, MUC5AC and MUC6 and de novo expression of MUC2.58
Bacterial interactions with mucins
Effects on mucins and their synthesis
Mucus is secreted by specialised goblet cells (also known as mucin-secreting cells).59–61 Through the expression of different mucins, variations in mucin glycosylation and the co-secretion of mucin-associated molecules, the nature of the mucus varies at different anatomical locations and constitutes a responsive system that can be adapted to local physiological requirements, including response to bacterial colonisation and infection.7–9 15 60 The mucins are produced by cells in the epithelial surface and/or by glands located in the submucosal connective tissues, and secretion occurs via both constitutive and regulated pathways.8 15 The constitutive pathway continuously secretes sufficient mucin to maintain the mucus layer, whereas the regulated pathway affords a massive discharge of mucus in response to environmental and/or pathophysiological stimuli such as bacterial infection and inflammation. Although release is regulated by activation of signalling pathways by a number of secretagogues (including acetylcholine, vasoactive intestinal peptide and neurotensin), activation can occur by bacterial-induced inflammatory mediators (eg, interleukin (IL)-1, IL-4, IL-6, IL-9, IL-13, interferons, tumour necrosis factor α and nitric oxide).7 8 59 61 The responsiveness to cytokines and other immune regulators provides a link between mucin-based defence, innate mucosal immunity and mucosal inflammatory responses.8 Stimulated mucin release occurs rapidly and is accompanied by a hundredfold or so expansion of secretory granules.
Changes in goblet cell function and in the chemical composition of intestinal mucins have been detected in response to a broad range of luminal insults, including changes in the normal microbiota and intrusion of enteric pathogens.41 42 Probiotic bacteria have been shown to be capable of enhancing mucin secretion in colonic epithelial cells,62 thus potentially boosting this defensive mechanism against pathogens. There is evidence that a variety of microbial products can stimulate increased production of mucins by mucosal epithelial cells.61–63 Also, germ-free mice exhibit changes in mucin gene expression, mucus composition and mucus secretion in response to intestinal bacteria or host-derived inflammatory mediators.64 65 Therefore, it can be surmised that alterations in mucus secretion and composition, including mucin glycosylation, represent important mechanisms for unfavourably changing the niche occupied by a mucosal pathogen. Successful pathogens, however, are capable of countering this mechanism. For example, H pylori has been shown to decrease mucin exocytosis,66 and LPS of this bacterium has been shown to affect mucin synthesis67 and the macromolecular nature of the mucin secreted,68 thereby representing a potential strategy by which the bacterium can favourably modulate mucin composition of the mucus barrier and the glycocalyx.
Specifically, H pylori causes an aberrant expression of human MUC1, MUC5AC and MUC6 in infected gastric mucosa,69 70 in particular with downregulation of MUC5AC and upregulation of MUC6,71 both of which are secreted mucins. A similar downregulation of MUC5AC has been found in rhesus monkeys, which have strong similarities to mucin glycosylation in humans.72 The reduced MUC5AC synthesis induced by H pylori can impair the gastric mucus gel,9 but the upregulation of MUC6 may represent an antimicrobial host response, since the O-linked glycans of MUC6 display antimicrobial activity against H pylori.73 The basis for this is the inhibition of biosynthesis of the essential bacterial membrane lipid, cholesterol-α-d-glucopyranoside, by the terminal α(1→4)-linked GlcNAc of MUC6 interacting with the respective enzyme.74 A close association with secreted MUC5AC and membrane-bound MUC1 is considered important for H pylori colonisation of the gastric mucosa.75 76 H pylori is bound by secreted MUC5AC through the bacterial outer membrane protein, BabA, by recognising fucosylated Leb expressed in this mucin.49 Although, on the one hand, aiding H pylori colonisation of the mucus layer, this binding may, on the other hand, limit colonisation to the mucus gel, and hence reduce the number of H pylori bacteria interacting with the epithelial surface by blocking BabA recognition of structures in the epithelial glycocalyx. Nevertheless, H pylori is capable of binding cell surface MUC1,75 77 and thus bacterial contact with the epithelial surface can be mediated in a mucin-dependent manner.
Other bacterial effects on mucins
In addition to direct interactions with mucins and their synthesis, bacteria can use other strategies to overcome the gel-based barrier function of the mucus layer. Flagellation is a shared characteristic of the vast majority of successful mucosal pathogens that are highly motile, such as C jejuni and H pylori.78 Motility, along with the corkscrew morphology of these bacteria, allows them to burrow through the mucus layer to reach the epithelium.79 Indeed, H pylori that have dysfunctional flagella have a greatly reduced ability to infect.80 Combined with motility, degradative enzymes, such as fucosidases, mucinases (mucin-degrading enzymes), glycosylsulfatases, sialidases, sialate O-acetylesterases and N-acetylneuraminate lyases, are produced by a broad spectrum of bacterial pathogens that destabilise the mucus as well as removing mucin decoy carbohydrates for bacterial adhesins.9 11 12 81 The size of the mucus polymer is important for the formation of entangled gels and the viscous properties of mucus,8 15 and thus degradation of mucins will result in localised disintegration of mucus, thereby aiding bacterial penetration of the epithelium.
Supporting the view of the mucus gel as an effective infection barrier, avoidance of this layer has been suggested as a further strategy used by certain mucosal pathogens of the GI tract.8 Although M cells are only a very small percentage of mucosal epithelial cells, they are the major point of attachment and/or entry used by a large proportion of enteric bacteria, including C jejuni.82 Intestinal M cells, which capture and present microbial antigens to the underlying lymphoid tissue, do not produce gel-forming mucins and have a thin glycocalyx.83 M cells selectively express the sialyl-Lea antigen, whereas Lea is expressed on all epithelial cells of the GI tract.84 85 Thus, M cells with a different profile of glycocalyx glycoproteins84 85 represent a focal point of entry for mucosal pathogens. Once M cells are invaded or mucus is penetrated, a further strategy of GI tract pathogens is to disrupt the tight junctions between adjacent mucosal cells, thereby exposing the vulnerable lateral membranes not protected by the glycocalyx. Such a mechanism is used by H pylori, which affects the permeability of cellular tight junctions,86 penetrates intercellular junctions, and, mediated by a bacterial lectin and LPS, binds to the glycoprotein laminin in the extracellular matrix and basement membrane.28 87 The latter interaction has further significance since H pylori LPS inhibits recognition of laminin by an epithelial cell receptor (integrin), which has been hypothesised to contribute to lack of mucosal integrity.88
Cell surface mucins in protection
Although the cell surface mucins can act as recognition molecules for bacterial adhesins, these mucins may also show other protective effects against pathogenic bacteria. For example, MUC1 has been shown to protect p53-expressing epithelial cells in vitro from the effects of C jejuni cytolethal distending toxin, a genotoxin, and, as demonstrated in Muc1−/− mice, lowers gastric colonisation by C jejuni.89 It should be noted that glycocalyx mucins are capable of initiating intracellular signalling in response to bacteria, reflecting their reporter function, as well as their barrier role as constituents of the glycocalyx on the apical surface of mucosal epithelial cells.7 17 Importantly, the cytoplasmic domain of MUC1 can be cleaved, translocates to mitochondria, and, together with the p53 transcription factor, modulates the cell cycle and protects against the apoptotic response to genotoxic stress.90 91 This protective effect, first identified in cancer research, has been suggested to have evolved as part of the natural epithelial defence against microbial genotoxins.8
Another protective mechanism of cell surface mucins that has been hypothesised, again exemplified by MUC1, involves their role as releasable decoy ligands for mucosal pathogens attempting to anchor to the glycocalyx. Supportive evidence for such a role of MUC1 has been accumulated for both C jejuni89 and H pylori.77 Although H pylori can bind MUC1, primary murine gastric epithelial cells expressing MUC1 bind fewer H pylori than murine Muc−/− cells.77 These, at first apparently conflicting, results can be explained if MUC1 acts as a releasable decoy; the bacteria bind MUC1 on normal gastric epithelial cells, but this is then shed by the host92; however, owing to the absence of MUC1 on Muc1−/− murine cells, the bacteria attach to other epithelial receptors and hence are not lost. The findings are reflected in pathology, since Muc1−/− mice develop fivefold greater H pylori colonisation density, which is maintained for at least 2 months, whereas heterozygous mice have a lower level of gastric MUC1 expression and intermediate colonisation densities.77 Moreover, comparing H pylori-positive patients, those with MUC1 polymorphisms exhibiting short extracellular mucin domains are more likely to develop gastritis than those without,75 because the shorter MUC1 molecules will allow greater access of the bacterium to the cell surface. Likewise, infection experiments in Muc1−/− mice have shown greater susceptibility to infection, GI barrier transit, and pathology by C jejuni than in normal mice.89 Collectively, the results are consistent with the importance of MUC1 in shielding the epithelial surface from pathogen contact.
Mucin-associated molecules in bacterial infection
In addition to mucins, other mucosa-associated molecules that involve carbohydrate–ligand interactions can play an important role in determining the success of bacterial colonisation of epithelial surfaces.15 An example of such molecules is trefoil factors.9 These factors, comprising TFF1–3, are a group of small proteins involved in wound repair and protection of the integrity of the epithelial barrier; they are secreted by mucus-secreting cells and expressed in a site-specific manner in the GI tract.93 TFF1 segregates with MUC5AC in the gastric foveolar epithelium, TFF2 with MUC6 in the glandular epithelium in the distal stomach, whereas TFF3 and MUC2 are co-expressed by the goblet cells of the large and small intestine. The role of these factors in infection is exemplified in H pylori colonisation of the gastric mucosa. H pylori has been shown to bind avidly to dimeric TFF1, and this interaction enables binding to gastric mucin, suggesting that TFF1 acts as a linker molecule binding to both H pylori and MUC5AC.94 95 Although the H pylori–TFF1 interaction appears well-founded, recognition by TFF1 has been ascribed to Glc- and Man-related structures within H pylori LPS by in vitro testing.89 However, this requires further examination because the sugar Man is absent from this LPS,40 44 but is present in bacterial cell surface mannans which co-purify with LPS.96
Another group of molecules, the surfactant proteins, have relevance to antibody-independent pathogen recognition and clearance from the GI tract. Surfactant protein D (SP-D), a member of the collectin family, is a C-type lectin that is found in mucus-secreting cells within gastric pits and with maximal expression at the luminal surface.97 98 Levels of expression of SP-D are significantly increased in H pylori-associated antral gastritis compared with normal human gastric mucosa, and co-localises with infecting H pylori bacteria.96 SP-D−/− mice are more susceptible to Helicobacter colonisation, and exhibit decreased inflammation and immune response to infection, emphasising the important influence on the establishment of infection.99 Functioning as a lectin, SP-D binds H pylori by recognition of the carbohydrate-bearing O-chain of LPS, resulting in bacterial aggregation and immobilisation,97 100 thus aiding eventual phagocytosis and clearance. Nevertheless, to evade this important mechanism of innate immune recognition, escape variants within the infecting H pylori population can arise98 100 with modifications in their O-chain glycosylation, thus decreasing bacterial interaction with SP-D and aiding colonisation.100 This represents an important example of changes in bacterial glycosylation to overcome lectin-based microbial recognition in the GI mucosa.
Conclusions
GI glycosylation in health and disease
The GI tract and its mucosa is a complex ecosystem of resident microbiota, resident and recruited immune cells, and a responsive mucus layer and epithelium. In addition to the barrier function of the mucus gel and glycocalyx against pathogens, protection is augmented by the production of antimicrobial peptides, including defensins, by the host.7 101 For example, expression of the human cathelicidin LL-37102 and human β-defensin 2103 is upregulated in the gastric mucosa of H pylori-infected patients.
Not to be overlooked, the microbiota may influence host antimicrobial peptide production. Angiogenin-4, a molecule with bactericidal activity produced by Paneth cells, is induced by Bifidobacterium thetaiotaomicron, a dominant member of the gut microbiota.104 Also, Bifidobacterium cell wall proteins have been shown to induce β-defensin 2 production from human intestinal epithelial cells.7 Moreover, Gram-negative bacterial components, such as LPS, peptidoglycan105 and protein derived from flagella (FliC), can induce expression of human β-defensin 2,106 and hence is likely to be induced by resident intestinal colonisers. Although members of the resident microbiota have been described as inducing inhibition of other bacterial species, including pathogens, by the production of the antimicrobial compounds microcins and colicins,7 107 invading pathogens can also produce such compounds to shape the GI tract flora. H pylori produces a ceropin-like peptide, Hp(2-20),108 to which it is resistant, compared with other Gram-negative bacteria, because of modification of a membrane lipoglycan moiety.109
In general, the gut microbiota is no longer considered inert and stagnant but evolving, and influences the homoeostasis of the host,7 110 111 for example, by influencing the host immunity through the T regulatory cell response.110 111 The capsular polysaccharide A of the intestinal symbiont Bacteroides fragilis induces protection from experimental colitis, produced by Helicobacter hepaticus, through induction of tolerogenic T cells.112 Also, members of the microbiota can limit anti-inflammatory responses and colonisation,113 and establish a balanced relationship with the host.110 Overall, it has been deduced that commensal bacteria have evolved specialised molecules, such as polysaccharide A of B fragilis, to induce tolerogenic responses that coordinate tolerance of the micobiota in the gut, in contrast with pathogen colonisation.111 This represents another level of communication with the host that can be utilised by the microbiota.
By modifying GI glycosation, the gut microbiota can further influence this barrier against pathogens. Seminal work showed that, in germ-free rodents, the mucus layer is reduced by half and is less stable,114 and has an altered glycosylation profile compared with conventional hosts.18 41 The use of cDNA microarray analysis has shown the participation of a well-established intestinal microbiota in regulation of the gut mucus layer and composition.65 It has been suggested that the resident microbiota can affect goblet cell dynamics and mucin composition directly, by the release of bioactive factors, and indirectly by activating host cells.7 41 For example, Gram-negative LPS increases the expression of mRNA of MUC5AC and MUC5B and stimulates their secretion.61 A bacterial quorum-sensing signal molecule of Gram-negative bacteria may stimulate production of MUC5AC. A strain of Lactobacillus plantarum, a Gram-positive subdominant species of the microbiota, increases expression of MUC2 and MUC3 mRNA,115 which has been shown to inhibit the cell attachment of enteropathogenic E coli (EPEC) strains.116 Moreover, a cocktail of probiotic bacteria (Lactobacillus and Bifidobacterium spp.) has been shown to attenuate C jejuni association with, and translocation through, mucus-secreting intestinal cells in vitro.117
Therefore, in considering glycosylation as contributing to the structures and barrier functions of the GI tract against pathogens, the influence of the resident microbiota promises to be a subject of developing interest and potential.
Glycosylation and glycosylated receptors as novel therapeutic targets
Biomedical applications of glycobiology have focused greatly on the roles of carbohydrates as receptors for adhesion of pathogens and toxins to host cells, appearance as tumour-associated markers, the regulation of inflammation, and the circulatory half-life of plasma glycoproteins and glycosylated therapeutic proteins. Many approaches have been adopted to modulate biological systems involving carbohydrates and lectin interactions. The most common involve the utilisation of glycans or glycomimetics to regulate the activity of lectins. The other approaches include modulation of genes and enzymes involved in glycosylation pathways and the use of glycotopes as vaccination targets.118 The success of sialic acid analogues (zanamivir and oseltamivir—Relenza and Tamiflu, respectively) as inhibitors of influenza virus sialidase shows promise for the use of glycan analogues against pathogens.119
The increasing antibiotic resistance associated with H pylori eradication by antimicrobial therapy is raising the need to search for alternative strategies. As H pylori uses the host glycan receptors for adhesion and infection, a strategy of carbohydrate-dependent inhibition of H pylori colonisation using carbohydrate-binding mimics (bio-mimics) will have a therapeutic and/or prophylactic effect on H pylori infection. Although H pylori-induced changes in host glycosylation have been implicated in the progression of gastric disease, the molecular mechanisms by which H pylori modulates host cell glycosylation is not known. In addition, the potential of using synthetic carbohydrates to inhibit H pylori adhesion to glycan receptors on gastric epithelial cells needs to be explored. Characterisation of the glycosylation genes and glycan receptors essential for H pylori adhesion will allow identification of individuals at highest risk of developing H pylori infection-associated diseases, as well as provide the proof of concept towards the design of new infection eradication strategies based on inhibition of bacterial adhesion to gastric epithelial cells.
Key points
All cells, including those of the gastrointestinal tract, are covered in a glycocalyx composed of glycolipids, glycoproteins and proteoglycans.
Mucins are a major class of glycoproteins that are both cell-membrane-bound and secreted to form a gel.
Mucins are key molecules in acute and chronic infections of the gut, and their glycosylation changes in response to microbial pathogens.
The supramucosal gel is a physicochemical sensor responsive to the environment that acts as a highly hydrated, mechanical and lubricative barrier and a matrix for immune-related and antimicrobial proteins.
Bacterial surface proteins, adhesins, bind to carbohydrate moieties on host cells and affect the adhesion and colonisation process.
Glycan–lectin interactions are emerging as key elements in gut pathogenesis.
The glycocalyx and carbohydrate components of the mucus gel are responsive to pathogens, which can promote or hinder adhesion and infection.
Acknowledgments
It is with deep sadness that we acknowledge that Professor Anthony Moran passed away on 30 September 2010. We dedicate this review to his memory.
References
Footnotes
Funding The authors were supported by Science Foundation Ireland grant No 08/SRC/B1393.
Competing interests None.
Provenance and peer review Commissioned; externally peer reviewed.