Article Text

Abstract
Objective Alcoholic liver disease (ALD) is a global health problem with limited therapeutic options. Intestinal barrier integrity and the microbiota modulate susceptibility to ALD. Akkermansia muciniphila, a Gram-negative intestinal commensal, promotes barrier function partly by enhancing mucus production. The aim of this study was to investigate microbial alterations in ALD and to define the impact of A. muciniphila administration on the course of ALD.
Design The intestinal microbiota was analysed in an unbiased approach by 16S ribosomal DNA (rDNA) sequencing in a Lieber-DeCarli ALD mouse model, and faecal A. muciniphila abundance was determined in a cohort of patients with alcoholic steatohepatitis (ASH). The impact of A. muciniphila on the development of experimental acute and chronic ALD was determined in a preventive and therapeutic setting, and intestinal barrier integrity was analysed.
Results Patients with ASH exhibited a decreased abundance of faecal A. muciniphila when compared with healthy controls that indirectly correlated with hepatic disease severity. Ethanol feeding of wild-type mice resulted in a prominent decline in A. muciniphila abundance. Ethanol-induced intestinal A. muciniphila depletion could be restored by oral A. muciniphila supplementation. Furthermore, A. muciniphila administration when performed in a preventive setting decreased hepatic injury, steatosis and neutrophil infiltration. A. muciniphila also protected against ethanol-induced gut leakiness, enhanced mucus thickness and tight-junction expression. In already established ALD, A. muciniphila used therapeutically ameliorated hepatic injury and neutrophil infiltration.
Conclusion Ethanol exposure diminishes intestinal A. muciniphila abundance in both mice and humans and can be recovered in experimental ALD by oral supplementation. A. muciniphila promotes intestinal barrier integrity and ameliorates experimental ALD. Our data suggest that patients with ALD might benefit from A. muciniphila supplementation.
- Akkermansia muciniphila
- alcoholic steatohepatitis
- alcoholic liver disease
- intestinal microbiota
- gut barrier
Statistics from Altmetric.com
- Akkermansia muciniphila
- alcoholic steatohepatitis
- alcoholic liver disease
- intestinal microbiota
- gut barrier
Significance of this study
What is already known on this subject?
The development of alcoholic liver disease (ALD) is strongly influenced by the microbiota.
Disruption of intestinal barrier function may promote ALD.
Akkermansia muciniphila supports maintenance of intestinal barrier integrity.
What are the new findings?
A. muciniphila abundance is diminished in patients with alcoholic steatohepatitis.
Ethanol exposure depletes intestinal A. muciniphila abundance in wild-type mice.
A. muciniphila supplementation ameliorates experimental ALD in a preventive and therapeutic setting.
Ethanol-induced disruption of intestinal barrier integrity is restored by A. muciniphila.
How might it impact on clinical practice in the foreseeable future?
Our data suggest that oral supplementation of A. muciniphila is a promising treatment strategy in human ALD.
Introduction
Alcoholic liver disease (ALD) is the most common cause for liver-related deaths worldwide and is responsible for 5.9% of all global deaths.1 2 ALD encompasses simple steatosis, fibrosis and cirrhosis all of which can deteriorate towards acute alcoholic steatohepatitis (ASH) with high mortality rates.3 Despite our increasing understanding of ALD pathogenesis, treatment strategies remain scarce.4
Disease processes in ALD are multilayered and involve direct ethanol-toxic effects on hepatocytes (eg, emergence of reactive oxygen species), but also indirect inflammatory signalling via pathogen-associated molecular patterns (PAMPs; eg, lipopolysaccharides (LPSs)) mainly derived from the intestine.5 6 These PAMPs potently activate hepatic macrophages (Kupffer cells) and promote an inflammatory response driven by interleukin 1 beta (IL-1β), interleukin 8 and tumour necrosis factor alpha (TNF-α). These cytokines consequently attract leucocytes, which promote hepatic injury.7–10 A similar situation appears to occur in human ALD, as patients exhibit a tight correlation between endotoxaemia and the degree of liver injury.11–13 Translocation of PAMPs may be facilitated by ethanol and ethanol-derived metabolites (eg, acetaldehyde) as a consequence of the disruption of tight junctions14 15 and intestinal compartmentalisation.16 Furthermore, evidence accumulates that the microbiota modulates the susceptibility to ALD.13 17 18 In line with this, ethanol-associated dysbiosis promotes hepatic disease and Lactobacillus spp ameliorate experimental ALD.19–21
Akkermansia muciniphila is a Gram-negative anaerobic commensal that utilises host-derived mucins as carbon and nitrogen source.22 In healthy individuals, A. muciniphila constitutes 1% to 4% of the faecal microbiota.23 24 A. muciniphila promotes mucus thickening and intestinal barrier function (ie, reduction of systemic endotoxin concentration) in mice on a high-fat diet.25 26 We aimed to determine the impact of ethanol consumption on A. muciniphila abundance in mice and humans and the impact of A. muciniphila in the development of experimental ALD.
Material and methods
Human studies
A. muciniphila abundance was quantified in stool samples of patients with ALD (ASH, n=21, age 50.9 years±10.04 or severe ASH, n=15; age 55.1 years±11.95)20 and in non-obese healthy individuals (n=16; age 41.1 years±2.6). Alcoholic patients were characterised by (1) a daily alcohol consumption of 50 g ethanol over the last year and (2) a liver biopsy with presence of acidophilic bodies, ballooning of hepatocytes, Mallory bodies, neutrophil infiltration, steatosis and fibrosis. Liver biopsies were scored by two independent, blinded observers for features of alcoholic hepatitis (ballooning degenerations, necrotic areas and infiltrated polymorphonuclear cell (PMNs). A widely used scoring system27 was used to assess acidophilic bodies (0: none; 1: ≤1/lobule or nodule; 2: >1/lobule or nodule), clarification/ballooning of hepatocytes (0: none; 1: not in all lobules/nodules; 2: in all lobules/nodules), Mallory bodies (0: none; 1: ≤1/lobule or nodule; 2: ≥2/lobule or nodule), neutrophil infiltrate (0: none; 1: isolated or rare; 2: marked). Steatosis was classified into five grades: 0: none; 1: mild (1% to 5% of hepatocytes); 2: moderate (6% to 32%); 3: marked (33% to 66%); and 4: severe (67% to 100%). Fibrosis was determined by a modified METAVIR score27: 0: no fibrosis; 1: pericentral and/or periportal fibrosis without fibrous septa; 2: pericentral and/or periportal fibrosis with few fibrous septa; 3: many fibrous septa without cirrhosis; and 4: cirrhosis. Patients were classified into a (non-severe) ASH group: score <6 and >2 or score ≥6 but without neutrophil infiltrate; and severe ASH: score ≥6 and neutrophil infiltrate score ≥1. Based on these histological features, patients were rated as ASH or severe ASH as described previously.20 Patient’s stool samples were collected during their hospitalisation at a single centre in France, and immediately frozen and stored at −80°C.
Mouse experiments
Four different models of experimental ALD were used to study ethanol-induced microbial changes and the efficacy of A. muciniphila supplementation. All experiments were aligned to ethical principles according to legal laws. (1) To study the influence of ethanol on microbiota composition, C57BL/6 wild-type (WT) mice were treated with a 10-day acute-on-chronic alcohol feeding model described previously.28 These experiments were performed in the Szabo laboratory. (2) To study the influence of A. muciniphila supplementation in ALD, 7- to 8-week-old female WT mice were fed a Lieber-DeCarli diet28–30 containing 1–5 vol% (ethanol fed) ad libitum for 15 days. Ethanol-fed or pair-fed mice were treated with A. muciniphila (1.5×109 colony-forming units (CFU)/200 µl phosphate-buffered saline (PBS)) or vehicle (PBS) by intragastric infusion with a 24-gauge stainless steel free tube every other day, starting on day 1. (3) To investigate the therapeutic properties of A. muciniphila in ALD, mice were fed with a Lieber-DeCarli diet for 15 days. After liver injury was verified by alanine transaminase (ALT) measurement on day 9, A. muciniphila administration (1.5×109 CFU/200 µl PBS) was carried out only on days 10, 12 and 14. (4) In the acute model of ethanol toxicity, mice were treated with A. muciniphila (1.5×109 CFU/200 µl PBS) on 2 days prior to the gavage of 6 g ethanol per kilogram bodyweight. Eight hours after ethanol gavage mice were sacrificed. All animals were anaesthetised before exsanguination and tissue sampling. In models 2–4, mice were harvested between 08:00 and 11:00.31
16S sequencing and bioinformatics
Caecal stool was collected directly from extracted caecum and frozen at −80°C. DNA was extracted using Stool DNA Extraction Kit (Qiagen) according to the manufacturer’s recommendations. Sequencing as described previously by Fadrosh et al 32 was completed at the Cincinnati University, Children’s Hospital Medical Center. Samples were excluded from downstream analysis based on quality measures determined by the sequencing facility, and one sample each from the ethanol-fed and pair-fed groups was excluded based on insufficient sequencing data. Sequence data were processed by UPARSE,33 and UTAX34 was used to generate operational taxonomic unit tables from 16S ribosomal DNA (rDNA) read data and to make taxonomic assignments. Linear discriminant analysis effect size (LEfSe)35 was applied to process 16S sequence data. LEfSe differences among biological groups were tested for significance using a non-parametric factorial Kruskal-Wallis sum-rank test followed by Wilcoxon rank-sum test. 16S rDNA sequencing data have been deposited in the National Center for Biotechnology Information GenBank repository under accession numbers: KY571432–KY572675.
Quantification of A. muciniphila in patient’s stool
Bacterial DNA was obtained by homogenising stools in a guanidinium thiocyanate lysis buffer using a Fast Prep homogeniser. High-quality bacterial DNA was extracted by successive steps of purification and precipitation.36 The primers used to detect A. muciniphila were based on 16S rDNA gene sequences: forward CAG CAC GTG AAG GTG GGG AC and reverse CCT TGC GGT TGG CTT CAG AT.25 Detection was achieved with a Light Cycler 480 (Roche Diagnostics, Basel, Switzerland) using the LC FastStart DNA Master SYBR Green I kit (Roche Diagnostics) with a primer concentration of 10 µM and annealing temperature of 60°C. The cycle threshold of each sample was then compared with a standard curve, performed in duplicate, made by diluting genomic DNA from A. muciniphila (DSM 22959) purchased from DSMZ. The data are expressed as nanogram of A. muciniphila DNA per gram of faecal content.
Cultivation of A. muciniphila
Culture of A. muciniphila MucT (CCUG 64013) was obtained from CCUG (Culture Collection, University of Göteborg, Sweden) and grown on chocolate agar (Biomerieux, Marcy l’Etoile, France) under anaerobic conditions at 37°C. For maintaining the anaerobic conditions GENbox and GENbox anaer systems (Biomeriux) were used. Before the administration, A. muciniphila were scraped from the agar plates, diluted in sterile PBS and were kept under anaerobic conditions until administration.26 Every mouse was orally administered with 1.5×109 CFU A. muciniphila. As control, sterile PBS was used.
Statistical analysis
For analysing our data, we used GraphPad PRISM 5 (La Jolla, California, USA). Unpaired two-tailed Student’s t-test, Kruskal-Wallis test followed by Dunn’s multiple comparison test, one-way analysis of variance followed by post hoc Newman-Keuls test and Spearman correlation test were used where appropriate. Two or more independent experiments were performed for each modality. Results are shown as mean±SEM. Statistical significance was considered at p<0.05.
Further information on materials and methods are provided in the online supplementary material.
Supplementary material 1
Results
Ethanol depletes A. muciniphila abundance
As the intestinal microbiota modulates susceptibility to ALD,13 18 37 38 we aimed at identifying bacterial species that would impact on ethanol-induced hepatic disease processes in an unbiased fashion. To do so, we exposed mice to a Lieber-DeCarli diet containing 5% ethanol and performed 16S rDNA sequencing on caecal content. We noted by LEfSe that ethanol feeding prominently reduced or increased the abundance of numerous taxonomic groups (figure 1A, online supplementary figure 1A). The cladogram in figure 1A shows those taxonomies that are significantly enriched in pair-fed animals (green) or in ethanol-fed mice (red) (‘p_’, phylum; ‘c_’, class; ‘f_’, family; ‘o_’, order; ‘g_’, genus). A. muciniphila and all higher taxonomies including its phylum Verrucomicrobia were significantly enriched in pair-fed animals (ie, reduced by ethanol feeding). We confirmed that A. muciniphila was enriched in pair-fed mice by quantitative PCR from caecal faeces and stool (figure 1B, online supplementary figure 1B). Ethanol stimulation, however, did not limit A. muciniphila growth in vitro (see online supplementary figure 1C).

Chronic alcohol consumption reduces faecal Akkermansia muciniphila numbers. (A) Analysis of taxonomic abundances using LEfSe indicates that multiple taxa, including A. muciniphila, are differentially enriched in the caecal content of control or ethanol-fed animals. The taxonomic groups indicated at right are differentially enriched in the corresponding group: red (ethanol fed) and green (pair fed). (B) Quantification of A. muciniphila numbers in caecal content measured by qPCR. (C) Quantification of A. muciniphila numbers in patients with ASH (n=21) and severe ASH (n=15) compared with healthy controls (n=16). (D) Correlation of ASH severity with A. muciniphila numbers. Data are shown as mean±SEM, n: (A, B) EtOH fed=9; pair fed=8. *p<0.05; **p<0.01; ***p<0.001 according to Kruskal-Wallis test followed by Wilcoxon rank-sum test (A), two-tailed Student’s t-test (B), one-way ANOVA followed by Newman-Keuls multiple comparison (C) and Spearman correlation (D). AH, alcoholic hepatitis; ANOVA, analysis of variance; ASH, alcoholic steatohepatitis; EtOH, ethanol; LEfSe, linear discriminant analysis effect size; qPCR, quantitative PCR; sAH, severe alcoholic hepatitis.
We then turned to our histologically confirmed human ASH cohort (see online supplementary tables 1 and 2) and analysed faecal A. muciniphila concentrations. We found reduced numbers of faecal A. muciniphila in patients with ASH when compared with healthy controls (figure 1C). A. muciniphila abundance showed a significant indirect correlation (r=−0.33, p=0018) with histological disease severity (figure 1D). Furthermore, we correlated faecal A. muciniphila counts with clinical parameters and observed a negative correlation between A. muciniphila and fibrosis (r=−0.5, p=0.01)39 and a positive correlation with high-density lipoprotein and low-density lipoprotein (see online supplementary table 3).
A. muciniphila supplementation protects against experimental ALD
To assess the impact of reduced A. muciniphila abundance on the development of ALD, we challenged WT mice that were either pretreated with A. muciniphila or vehicle (PBS) by oral gavage with a single shot of ethanol (figure 2A). PBS-treated WT mice developed hepatic injury and inflammation, indicated by ALT elevation (figure 2B), IL-1β expression (figure 2C) and neutrophil infiltration (figure 2D,E), respectively. In contrast, A. muciniphila pretreated WT mice were protected against acute ethanol-induced hepatic disease (figure 2B–E).

Acute ethanol-induced injury was prevented by Akkermansia muciniphila supplementation. (A) Experimental design; arrows indicate A. muciniphila administration. (B) Serum ALT concentrations. (C) IL-1β expression normalised to β-actin determined by qPCR. (D and E) Representative images and quantification of MPO+ cells per high power field in the liver based on hepatic MPO immunoreactivity (brown, indicated by black arrows). Data are shown as mean±SEM, n: (B–E) EtOH=11, EtOH+A.muc=12, Ctrl=5, Ctrl+A.muc=6. *p<0.05; **p<0.01; ***p<0.001 according to one-way ANOVA followed by Newman-Keuls multiple comparison (B–E). ALT, alanine transaminase; A.muc, Akkermansia muciniphila; ANOVA, analysis of variance; EtOH, ethanol; HPF, high power field; IL-1β, interleukin 1 beta; MPO, myeloperoxidase; PBS, phosphate-buffered saline; qPCR, quantitative PCR.
In a next step, we assessed whether supplementation of A. muciniphila protected against ALD in a chronic disease model. We exposed mice to a Lieber-DeCarli diet containing 5% ethanol for 15 days and gavaged A. muciniphila every other day (figure 3A). Supplementation restored A. muciniphila abundance in ethanol-fed WT mice (figure 3B). Vehicle-treated ethanol-fed WT mice exhibited signs of hepatic injury, with significantly increased liver-to-body-weight ratio (figure 3C), elevated ALT levels (figure 3D) and significantly increased expression of proinflammatory cytokines such as TNF-α and IL-1β (see online supplementary figure 2A,B). Furthermore, vehicle-treated ethanol-fed WT mice exhibited hepatic inflammation indicated by myeloperoxidase-positive (MPO+) neutrophilic infiltration (figure 3E,F), and steatosis (figure 3G,H, online supplementary figure 2C). Importantly, A. muciniphila-treated mice were protected against hepatic injury (figure 3C,D), infiltration of MPO+ neutrophils (figure 3E,F) and steatosis (figure 3G,H). Expression of inflammatory cytokines were insignificantly decreased (see online supplementary figure 2A,B,D–E). The Lieber-DeCarli 5% ethanol diet did not induce hepatocyte ballooning, necrotic bodies (data not shown) or Kupffer cell activation (see online supplementary figure 2F).

Treatment with Akkermansia muciniphila protected against experimental ALD. (A) Experimental design; arrows indicate A. muciniphila administration. (B) A. muciniphila abundance determined by qPCR. (C) Liver-to-body-weight ratio and (D) serum ALT concentration. (E and F) Representative images and quantification of MPO+ cells per high power field in the liver based on hepatic MPO immunoreactivity (brown, indicated by black arrows). (G and H) Biochemical and histological assessment of steatosis with representative pictures of oil-red-O stained liver sections. Data are shown as mean±SEM, n: (B–F) EtOH fed=10, EtOH fed+A.muc=10, pair fed=5, pair fed+A.muc=4. (G–H) EtOH fed=13, EtOH fed+A.muc=10, pair fed=5, pair fed+A.muc=6. *p<0.05; **p<0.01; ***p<0.001 according to one-way ANOVA followed by Newman-Keuls multiple comparison test (B–H). ALD, alcoholic liver disease; ALT, alanine transaminase; A.muc, Akkermansia muciniphila; ANOVA, analysis of variance; EtOH, ethanol; HPF, high power field; MPO, myeloperoxidase; qPCR, quantitative PCR.
Supplementation with A. muciniphila ameliorates established ALD
Next, we tested whether A. muciniphila treatment could be therapeutically used in already established ALD. To do so, WT mice were exposed to a Lieber-DeCarli diet for 15 days, and A. muciniphila treatment was started on day 10 (figure 4A). By this time, ethanol-induced hepatic injury was clearly detectable and comparable in both treatment groups (figure 4B). Notably, oral gavage of A. muciniphila for 3 days ameliorated hepatic injury in WT mice, indicated by reduced ALT levels (figure 4C). In line with this, A. muciniphila-treated mice exhibited reduced hepatic expression of IL-1β (figure 4D), a trend towards reduced TNF-α expression (see online supplementary figure 3A), and significantly reduced infiltration of MPO+ neutrophils (figure 4E, F). Steatosis increased after ethanol feeding with a trend towards protection in 3-day A. muciniphila supplemented mice (figure 4G,H, see online supplementary figure 3B).

Akkermansia muciniphila ameliorated established ALD. (A) Experimental design; arrows indicate A. muciniphila administration. (B and C) Serum ALT concentration before (B) and after (C) A. muciniphila supplementation. (D) Expression of IL-1β normalised to β-actin determined by qPCR. (E and F) Representative images and quantification of MPO+ cells per high power field in the liver based on hepatic MPO immunoreactivity (brown, indicated by black arrows). (G and H) Histological assessment of steatosis with representative pictures of oil-red-O stained liver sections. Data are shown as mean±SEM, n: (B–D; G–H) EtOH fed=10, EtOH fed+A.muc=10, pair fed=4, pair fed+A.muc=4; (E–F) EtOH fed=15, EtOH fed+A.muc=10, pair fed=4, pair fed+A.muc=4. *p<0.05; **p<0.01; ***p<0.001 according to one-way ANOVA followed by Newman-Keuls multiple comparison (B–H). ALD, alcoholic liver disease; ALT, alanine transaminase; A.muc, Akkermansia muciniphila; ANOVA, analysis of variance; EtOH, ethanol; HPF, high power field; IL-1β, interleukin 1 beta; MPO, myeloperoxidase; qPCR, quantitative PCR.
A. muciniphila restores intestinal barrier function in ALD
We next explored putative mechanisms how A. muciniphila protected against ALD. We excluded the possibility that A. muciniphila metabolised ethanol in vitro (see online supplementary figure 4A). In line with this, we detected comparable ethanol concentrations in the blood from ethanol-fed WT mice with and without supplementation of A. muciniphila in all of our ALD models (see online supplementary figure 4B–D). We did also not find evidence that A. muciniphila treatment impacted on hepatic expression of ethanol-degrading enzymes (see online supplementary figure 4E–G). Furthermore, we were unable to detect relevant A. muciniphila DNA in the liver, suggesting that the protective effect may be originating from the intestine (see online supplementary figure 5H).
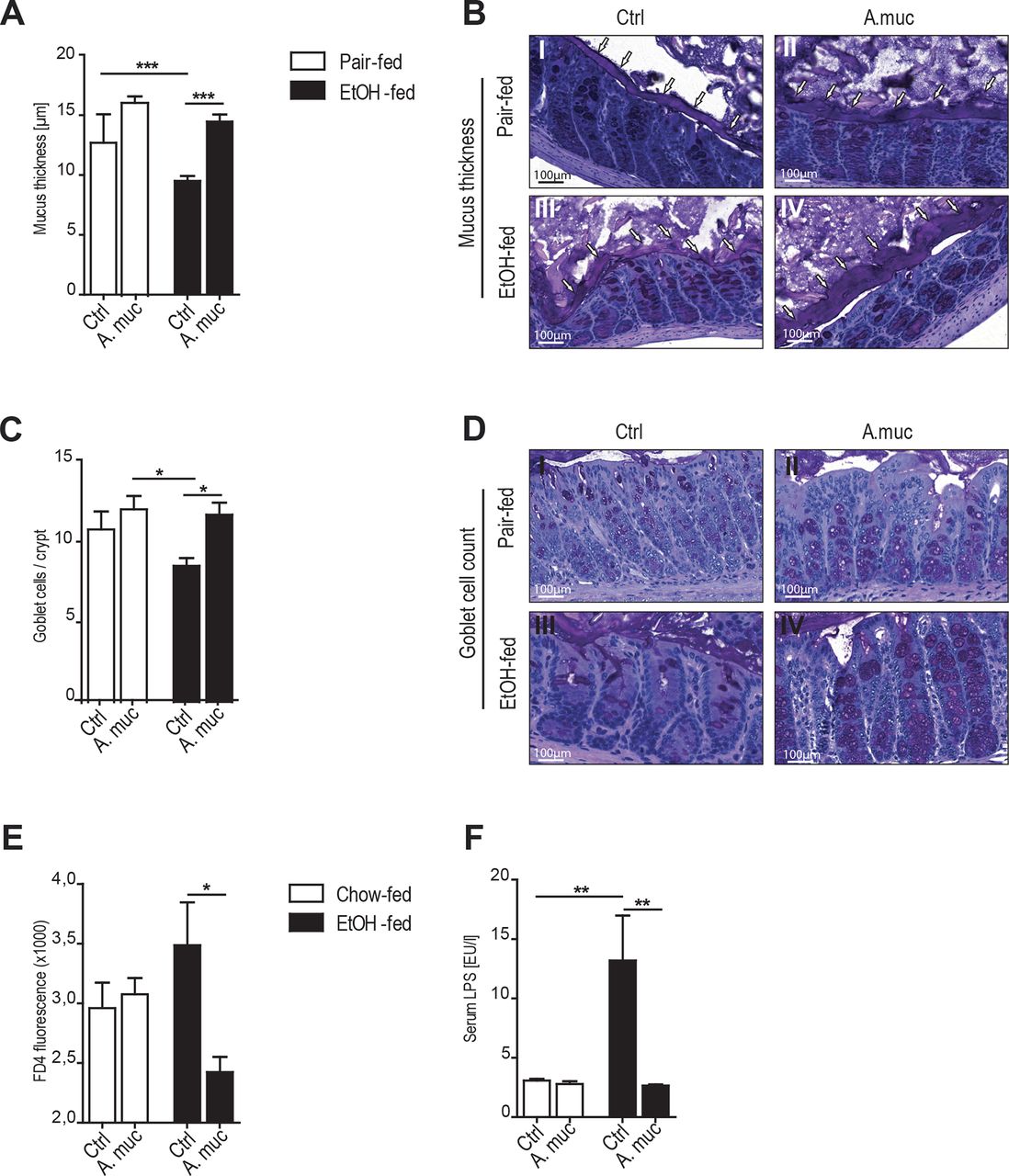
Oral supplementation of Akkermansia muciniphila restored intestinal barrier function. (A and B) Quantification of mucus thickness after 15 days of A. muciniphila supplementation in the chronic ALD model. (C and D) Quantification of colonic goblet cells identified by periodic acid–Schiff reaction. (E) Serum FD4 (fluorescein isothiocyanate dextran 4) level of the in vivo gut permeability assay. (F) Serum lipopolysaccharide concentrations in the chronic ALD model. Data are shown as mean±SEM, n: (A–E) EtOH fed=10, EtOH fed+A.muc=10, pair fed=5, pair fed+A.muc=4; (F) EtOH fed=20, EtOH fed+A.muc=11, pair fed=7, pair fed+A.muc=4. *p<0.05; **p<0.01; ***p<0.001 according to one-way ANOVA followed by Newman-Keuls multiple comparison (A–F). ALD, alcoholic liver disease; A.muc, Akkermansia muciniphila; ANOVA, analysis of variance; EtOH, ethanol; LPS, lipopolysaccharide.
Improved metabolic control by A. muciniphila in mouse models of obesity and type 2 diabetes was paralleled by increased mucus production at the epithelial surface and restored barrier function.25 26 40 As the intestinal barrier determines susceptibility to ALD,41–43 we assessed intestinal barrier integrity in ethanol-fed WT mice after 15 days with or without A. muciniphila supplementation. Ethanol feeding disrupted the mucus layer and diminished mucosal thickness compared with pair-fed WT mice (figure 5A,B). In contrast, treatment with A. muciniphila protected against ethanol-induced disruption of the mucus layer (figure 5A,B). This was associated with increased numbers of mucus-producing goblet cells in A. muciniphila-treated mice (figure 5C,D) and a thickened mucin 2 mucus layer, which appeared to be regulated post-transcriptionally (see online supplementary figure 5A–D).
To further investigate whether enhanced mucus production induced by A. muciniphila impacted on intestinal barrier function, we used a model in which fluorescein isothiocyanate (FD4) is exposed to the intestinal mucosa in vivo and tracked systemically in the serum to quantify gut leakiness. Intestinal A. muciniphila supplementation of ethanol-fed mice led to reduced systemic FD4 translocation compared with ethanol-fed controls (figure 5E), indicating that A.muciniphila indeed promoted mucosal barrier function.44 This was observed to a lesser extent when mice were exposed to a single acute ethanol shot (see online supplementary figure 5F). In line with a restored intestinal barrier function in ALD, we observed that A. muciniphila treatment reduced endotoxin levels in the serum of ethanol-fed mice (figure 5F) and tended to reduce LPS in our therapeutic approach (see online supplementary figure 5G). Systemic LPS concentration remained unaltered after a single ethanol shot (see online supplementary figure 5H).
Interepithelial tight junctions influence intestinal epithelial leakage and are critically important to maintain barrier integrity.45 46 As A. muciniphila protected against ethanol-induced gut leakiness, we assessed the expression of tight-junction proteins in colonic tissue. Ethanol feeding led to reduced expression of Claudin-3 and Occludin, which was restored by A. muciniphila treatment (figure 6A,B), while we did not observe any differences in Claudin-1 and Zona occludens-1 expression (data not shown). In line with this, we detected restored immunoreactivity of claudin-3 and occludin in colonic epithelial cells from A. muciniphila-treated mice compared with ethanol-fed controls (figure 6C–F).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Akkermansia muciniphila restores intraepithelial barrier function. (A) Fold expression of Cldn-3 and (B) Ocld in the colon tissue of mice exposed to 15-day ethanol and the Lieber-DeCarli diet. (C and D) Quantification of claudin 3 immunoreactivity (green) with representative confocal microscope images of murine colon. (E and F) Quantification of occludin immunoreactivity (red) with representative confocal microscope image of murine colon. DAPI, blue. Data are shown as mean±SEM, n: (A and B) EtOH fed=5, EtOH fed+A.muc=6, pair fed=5, pair fed+A.muc=5; (C–F) EtOH fed=3, EtOH fed+A.muc=6, pair fed=3, pair fed+A.muc=5. *p<0.05; **p<0.01; ***p<0.001 according to one-way ANOVA followed by Newman-Keuls multiple comparison (A–F). A.muc, Akkermansia muciniphila; ANOVA, analysis of variance; Cldn-3, claudin 3; DAPI, 4′,6-diamidino-2-phenylindole; EtOH, ethanol; Ocld, occludin.
Discussion
The intestinal microbiota has evolved as a major player in various hepatic disorders,47 and several studies have recently suggested that the gut microbiota is of major importance in ALD.48 Here, we demonstrated with an unbiased approach that ethanol consumption depletes intestinal abundance of A. muciniphila. In line with this, we noted reduced A. muciniphila abundance in patients with ASH, which indirectly correlated with hepatic disease severity. Oral supplementation of A. muciniphila recovered intestinal abundance and ameliorated ALD in an acute and chronic experimental setting.
In humans, alcohol intake has been associated with gut microbial alterations, and dysbiosis appears to be a driving force in ALD.6 11 13 18 20 Patients with ALD exhibit decreased abundance of Bacteroidetes and increased abundance of Proteobacteria.13 Llopis et al recently showed that the dysbiotic gut microbiota from patients with ASH promotes susceptibility to developing ALD.20 The dysbiosis in patients with alcoholic hepatitis was characterised by an increase in Bifidobacteria, Streptococci and Enterobacteria whereas Clostridium leptum and Faecalibacterium prausnitzii, both well-established anti-inflammatory strains,49 50 were decreased. Transfer of this dysbiotic human faecal microbiota into germ-free or conventionally raised mice conferred increased susceptibility to ALD.20 We now demonstrate that the commensal A. muciniphila (phylum Verrucomicrobia) is significantly diminished in patients with ASH and mice exposed to the Lieber-DeCarli ethanol diet. The latter observation was supported by 16S rDNA ribotyping and PCR quantification, respectively, in two different laboratories. In contrast, ALD studies using a different experimental approach demonstrated partly divergent results with increased A. muciniphila numbers after ethanol exposure.11 30 48 The mechanism by which ethanol depletes A. muciniphila abundance is currently not understood. As ethanol did not impair A. muciniphila growth in vitro, other (eg, indirect) mechanisms of A. muciniphila depletion may be in place which could involve microbial interactions in a dysbiotic community. Beside ethanol consumption, diet might directly influence intestinal A. muciniphila abundance; however, we were unable to collect dietary information in our ASH cohort, which represents a limitation of our study.
Recent reports revealed beneficial effects of A. muciniphila on host metabolism. Everard et al showed a significant reduction of A. muciniphila numbers in WT mice fed a high-fat diet and that supplementation of A. muciniphila improved metabolic dysregulation.25 51 The A. muciniphila membrane protein Amuc_1100 mediated this protective effect probably via modulation of TLR2 signalling.52 Similar to our study, the authors also demonstrated that A. muciniphila restored barrier function. Furthermore, in obese individuals A. muciniphila abundance negatively correlated with fasting glucose concentrations, waist-to-hip ratio and subcutaneous adipocyte diameter. In line with a protective effect of A. muciniphila on metabolism in obesity, patients with pronounced A. muciniphila abundance strongly benefited from calorie restriction.53 Li et al demonstrated that A. muciniphila supplementation in apolipoprotein E-deficient mice fed a western-diet protected against atherosclerosis.26 More specifically, A. muciniphila protected against systemic endotoxin mediated inflammation in atherosclerotic lesions, probably by restoration of intestinal barrier function. A. muciniphila also ameliorated acute and chronic hyperlipidaemia mediated by enhanced low-density lipoprotein receptor expression.54 In ALD, prebiotics (eg, the plant extract rhubarb) and Lactobacillus spp have been demonstrated to ameliorate hepatic inflammation and injury.55–59 However, our study is the first to directly demonstrate a protective effect of A. muciniphila in ALD. More specifically, A. muciniphila supplementation reduced ethanol-induced hepatic injury, steatosis and infiltration of MPO+ neutrophils, which critically drive ALD.60
A. muciniphila did not metabolise ethanol in vitro, and we observed comparable ethanol concentrations in the blood of A. muciniphila-treated mice compared with controls.21 We rather demonstrated that A. muciniphila protected against barrier disruption induced by ethanol exposure. Improved intestinal barrier function was exemplified by reduced gut leakiness, a restored expression of tight-junction proteins and a thickened mucus layer in A. muciniphila-treated WT mice. In line with this, systemic LPS level normalised in A. muciniphila-treated ethanol-exposed WT mice. As gut leakiness and LPS have been implicated in the development of ALD,9 17 our observations may provide one explanation how A. muciniphila protects against ALD. As A. muciniphila-treated mice were also protected from acute ethanol-induced injury that did not alter intestinal permeability in our hands, other mechanisms of A. muciniphila-mediated protection may be in place.
Based on these data, we propose that depletion of A. muciniphila reflects an early event in the pathophysiology of ALD probably by regulating gut barrier function. Recovery of ethanol-induced A. muciniphila depletion by oral supplementation could represent a novel treatment option for patients with ALD.
Acknowledgments
We thank Cindy Hugot for assistance with this study.
References
Footnotes
CG and TEA contributed equally.
Contributors CG designed, performed and analysed most experiments together with VW, FG, BE, LW, PL, DVW, BG, DC, SM, LM and AP. MD performed histology analysis, and AMC, PM, RRG, ARM, GP, GS and TEA provided expertise. CG and TEA prepared the manuscript. HT coordinated the project.
Funding This work was supported by Christian Doppler Research Society (to HT). HT is supported by the excellence initiative (Competence Centers for Excellent Technologies) of the Austrian Research Promotion Agency FFG: Research Center of Excellence in Vascular Ageing Tyrol, VASCage (K:Project Nr. 843536) funded by the Bundesministerium für Verkehr, Inovation und Technologie (BMVIT), Bundesministerium für Wissenschaft, Forschung und Wirtschaft (BMWFW), the Wirtschaftsagentur Wien and the Standortagentur Tirol. TEA is supported by the Austrian Science Fund (FWF) P 29379:B28 and the Tyrolian Science Fund (TWF) 0404/1812. GS and PL were supported by R01 AA017729 from the National Institute on Alcoholism and Alcohol Abuse, USA.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.