Article Text

Abstract
Background and aims Inappropriate immune responses contribute to the continuous stimulation of the intestinal immune system in chronic inflammatory bowel disease (IBD). Among several pathogenic factors, a numerical deficiency of regulatory T (Treg) cells has been suggested to lead to an insufficient compensation of chronically activated T lymphocytes. This study was conducted to investigate whether increased apoptosis contributes to Treg cell deficiency in IBD and whether successful treatment with antitumour necrosis factor α (TNFα) is achieved by reducing of Treg cell apoptosis.
Methods Apoptosis of CD4+Foxp3+ Treg cells in tissue sections of patients with active IBD was analysed by immunohistochemistry and TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labelling) staining. Apoptosis of peripheral blood CD4+CD25+Foxp3+ Treg cells was investigated by flow cytometry and annexin-V staining. In addition, caspase activity and apoptosis were measured in sera of patients with IBD treated with anti-TNFα by a luminometric caspase enzyme assay.
Results It is demonstrated that patients with active IBD revealed increased apoptosis of local CD4+Foxp3+ Treg cells in the inflamed mucosa compared with non-inflamed control colon tissue. Moreover, in peripheral blood a reduced frequency and increased apoptosis of Treg cells were found and accompanied by elevated caspase activity in the serum. During anti-TNFα treatment, Treg cell apoptosis declined in close correlation with elevated peripheral Treg cell numbers and a decrease of caspase activation and disease activity.
Conclusions These data suggest that increased apoptosis of Treg cells plays a potentially important role in the pathogenesis of IBD and can be reversed by anti-TNFα treatment. Measurement of Treg cell apoptosis and serum caspase activity might therefore represent promising tools for monitoring disease activity and treatment response in patients with IBD.
- Apoptosis
- inflammatory bowel disease
- Treg cells
- caspases
- anti-TNFα
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
Dysfunctions of immunosuppressive regulatory T (Treg) cells have been implicated in several autoimmune diseases. Inflammatory bowel disease (IBD) is characterised by a numerical deficiency of Treg cells, which was assumed to be caused by an enhanced outflow of peripheral Treg cells into the mucosa.
The molecular basis of the numerical deficiency of Treg cells remains unclear. It is also not known whether alterations of apoptosis and restoration of the Treg cell number are involved in the efficient treatment of patients with IBD with tumour necrosis factor α (TNFα) antagonists.
What are the new findings?
We show that apoptosis of Treg (CD4+Foxp3+) cells plays a potentially important role in the pathogenesis of IBD. The inflamed mucosa of patients with active IBD revealed a higher number but also increased apoptosis of local CD4+Foxp3+ Treg cells compared with non-inflamed mucosa of control colon tissue. Thus, patients with active IBD showed a deficiency in local Treg cells.
Patients with active IBD also revealed a reduced frequency of CD4+CD25+Foxp3+ Treg cells in peripheral blood, which was accompanied by increased apoptosis of Treg cells. IBD might therefore be characterised by a numerical deficiency rather than a functional defect of Treg cells in peripheral blood.
During anti-TNFα treatment, apoptosis of peripheral Treg cells and caspase activation declined in responders but not in non-responders, resulting in the restoration of peripheral Treg cell numbers and reduced disease activity.
How might it impact on clinical practice in the foreseeable future?
Increased apoptosis of peripheral and mucosal Treg cells might be involved in the pathogenesis of IBD and can be reversed by anti-TNFα treatment, thereby restoring T cell homeostasis. Measurement of Treg cell apoptosis and serum caspase activity might be promising tools for monitoring disease activity and treatment response in patients with IBD.
Introduction
In the normal gut system a balance between proinflammatory and anti-inflammatory responses guarantees the defence against pathogens and maintenance of tolerance to the commensal bacterial flora. A breakdown of the immunological tolerance leads to pathological inflammation and is thought to play an important role in chronic inflammatory bowel disease (IBD) including Crohn's disease (CD) and ulcerative colitis (UC).1 Data from animal and human studies showed that defects of the intestinal epithelial barrier and a dysregulated gut immune system can lead to IBD.2 3 In particular, an increased number of activated mucosal T lymphocytes is a typical finding in both CD and UC.4 The reasons for the enhanced immune response in IBD might be an excess of inflammatory mediators and an inadequate function or number of cellular components that downregulate mucosal immune responsiveness.
Among the key contributors to suppression of inappropriate immune responses are regulatory T (Treg) cells that normally constitute 1–5% of peripheral T cells.5–7 Treg cells were first recognised by their surface expression of CD4 and the interleukin 2 (IL-2) receptor-α chain/CD25 and are characterised by a high expression of transcription factor Foxp3, which is considered to confer their regulatory activity.8 9 Those cells exhibit potent suppressor activity by inhibiting the proliferation of effector T cells and the production of T helper 1 (Th1) and Th2 cytokines.10 A disrupted balance between effector T cells and Treg cells has been implicated in autoimmune diseases such as autoimmune hepatitis and multiple sclerosis.11 12 Treg cells are also essential for the maintenance of mucosal tolerance and capable of curing experimental colitis in animal models.13 14 So far, there are only a few studies describing the role of Tregs in human IBD, but increasing evidence indicates their involvement in the breakdown of immunological self-tolerance.15 16 A recent report demonstrated an increased frequency of functional Treg cells in mucosal lymphoid tissue in active IBD, which was however significantly lower compared with other inflammatory intestinal diseases.17 Moreover, the frequency of Treg cells was decreased in the peripheral blood during active disease, suggesting that a numerical rather than a functional defect of Treg activity might contribute to pathogenesis of IBD. The mechanisms underlying the numerical defect of Treg cells in patients with IBD remain unknown.
A key process for T cell homeostasis is apoptosis, which is controlled by intracellular proteases of the caspase family. Caspases cleave various cellular substrates including poly(ADP-ribose)polymerase (PARP) and thereby induce the typical alterations of apoptotic cells.18 Caspases can be activated by two signalling routes, namely the extrinsic and the intrinsic pathways. The intrinsic pathway is initiated by the mitochondrial release of cytochrome c, whereas the extrinsic pathway is triggered by death receptors binding the death ligands CD95L, tumour necrosis factor α (TNFα) or TNF-related apoptosis-inducing ligand (TRAIL).
Lamina propria T cells express both CD95 and CD95L and show enhanced susceptibility to CD95-mediated apoptosis,19 a feature that might represent a mechanism of regulating mucosal immune responses. Increased cytokine activation might also contribute to altered T cell apoptosis in IBD.20 For instance, increased expression of CD95L and TNFα has been demonstrated in patients with IBD. The central role of TNFα is underlined by the responsiveness of IBD to TNFα inhibition, such as treatment with infliximab.21 The mechanism by which TNFα regulates inflammation in IBD is likely to be complex. Although some studies indicate direct or indirect effects of TNFα on Treg cell function,22 23 it is unknown whether increased apoptosis of Treg cells contributes to IBD and how this process is influenced by TNFα inhibition.
In the present study, we investigated the role of Treg cell apoptosis in patients with active IBD and during the course of anti-TNFα treatment. Our data show that apoptosis is strongly elevated in mucosal and peripheral CD4+CD25highFoxp3+ Treg cells of patients with IBD. Interestingly, in patients treated successfully with anti-TNFα, the number of Treg cells was restored in close correlation with a decrease of Treg cell apoptosis and serological caspase activity. Our data suggest that increased Treg cell apoptosis is involved in IBD pathogenesis and can be reversed by anti-TNFα, thereby restoring Treg cell homeostasis.
Materials and methods
Patients
We investigated apoptosis of Treg cells in peripheral blood of patients with active UC (n=24, 14 men, 10 women, mean age 39.3±2.4 years, range 22–65 years) and active CD (n=32, 14 men, 18 women, mean age 36.4±2.1 years, range 20–62 years). Patients had no or only partial response to treatment with prednisolone and/or azathioprine. Healthy individuals (n=10, 5 men, 5 women, mean age 25.7±0.6 years, range 23–29 years) served as controls. Sera from 66 patients with active IBD (32 UC, 34 CD, 41 men, 25 women, mean age 37.8±1.5 years, range 20–69 years) and from 29 patients with IBD in remission (7 UC, 22 CD, 15 men, 14 women, mean age 35.3±2.5 years, range 19–62 years) were analysed for caspase activation. Eighteen healthy individuals served as controls (mean age 28±5 years, range 23–54 years). In 11 patients with active CD (2 men, 9 women, age 35.6±2.6 years, range 26–52 years) we analysed the number of Treg cells and serological caspase activity during anti-TNFα treatment. Patients received anti-TNFα (infliximab, 5 mg/kg) intravenously at weeks 0, 2 and 6, and then every 8 weeks in combination with azathioprine (2–2.5 mg/kg/day). Active CD was prospectively defined according to CDAI (Crohn's disease activity index) >150 and active UC according to Lichtiger score >3.24 25 The study was performed according to guidelines of the Ethics Committees of the Hannover Medical School and University of Heidelberg.
Immunohistochemical detection of caspase activation
Biopsies from colon or ileum of patients with active CD (n=5) and control colon (n=5) were obtained from the Department of Pathology of Hannover Medical School. Tumour-free colon tissues from patients with colon carcinoma served as controls. Rehydration, blocking of endogenous peroxidase and antigen retrieval of the paraffin sections was performed as described.26 Slides were incubated with activation-specific anti-caspase-3, anti-caspase-7 or anti-cleaved PARP, and then incubated with biotinylated secondary antibodies as described.27 Sections were covered with avidin–biotin complex reagent, stained in substrate solution (aminoethylcarbazole/hydrogen peroxide) and counterstained with haematoxylin.27
Immunohistochemical triple staining
Triple staining was performed for CD4, Foxp3 and TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labelling) reactivity. Sections (CD n=5, UC n=5, microscopic colitis n=4, controls n=5) were deparaffinised and antigens were retrieved by boiling for 5 min in antigen unmasking solution (Vector Laboratories, Burlingame, CA, USA). After blocking non-specific binding with 4% bovine serum albumin (BSA), sections were incubated overnight at 4°C with mouse anti-CD4 (1:25, Abcam, Cambridge, Massachusetts, USA). After washing in phosphate-buffered saline (PBS), sections were incubated with Cy5-conjugated goat antimouse antibody (1:200, Dianova, Hamburg, Germany) for 30 min, and then permeabilised with 0.1% Triton X-100 in 0.1% sodium citrate. Sections were washed again and incubated for 1 h with biotinylated mouse anti-Foxp3 (clone PCH101, 1:50, eBioscience, San Diego, California, USA). After repeated washings, sections were incubated with streptavidin–Cy3 conjugate for 30 min (1:1000, Dianova). They were then subjected to TUNEL staining using the in-situ cell death detection kit (Roche, Penzberg, Germany) and incubated for 1 h at 37°C in a reaction mixture containing terminal deoxynucleotidyl transferase and fluorescein isothiocyanate (FITC)-labelled dUTP. After 4′,6-diamidino-2-phenylindole (DAPI) staining, CD4+Foxp3+ cells with or without TUNEL reactivity were counted in four microscopic fields at ×400 magnification.
Serological detection of caspase activation
Caspase-3 and -7 activities in serum samples were measured as described using a proluminescent substrate coupled to the tetrapeptide DEVD.26 In addition, sera were measured for the caspase-generated cytokeratin-18 fragment using the M30 ELISA (Peviva, Bromma, Sweden).28
Flow cytometric detection of apoptosis of Treg cells
Peripheral blood mononuclear cells were isolated on a Ficoll-Paque gradient and seeded in 96-well plates (2×106/well). Cells were incubated with PerCP-conjugated anti-CD4 and APC-conjugated anti-CD25 (10 μg/well, Becton Dickinson, Heidelberg, Germany) for 30 min. After washing in PBS/1% BSA, cells were incubated with FITC-conjugated annexin-V (Roche), treated with permeabilisation buffer (eBiosciences) and stained with phycoerythrin (PE)-conjugated anti-Foxp3 (clone PCH101) for 1 h. After washing with the buffer of the permeabilisation kit, the percentage of CD4+CD25highFoxp3+ cells among the CD4-positive T cells and the percentage of annexin-V-positive cells in the CD4+CD25highFoxp3+ population were quantified in a FACS Calibur flow cytometer (Becton Dickinson). Expression of surface markers was assessed after gating on lymphocytes by scatter characteristics. Unstained cells and cells stained with isotype-matched control antibodies served as controls.
Statistical analysis
A statistical analysis comparing results from healthy individuals and patients with IBD was performed by using the two-tailed unpaired (figures 3–5) or paired (figure 7) t test for equality of means (SPSS 17.0 software). The statistical analysis was confirmed by a professional statistician. A p value of <0.05 was considered significant.
Results
In situ detection of caspase activation and PARP cleavage in patients with IBD
To investigate the role of apoptosis in IBD, we first analysed the activation of effector caspases, such as caspase-3 and caspase-7, in biopsies using activation-specific antibodies against both caspases. In addition, we assessed the cleavage of the caspase substrate PARP using a cleavage-specific antibody. Almost no immunoreactivity with all three antibodies was detected in control colon tissue (figure 1A–C), indicating a lack of apoptosis in these biopsies. In contrast, when analysing inflamed tissues from five patients with active CD, we consistently found many lamina propria cells that stained positively for active caspase-3 and -7 as well as for cleaved PARP (figure 1D–F). Active caspase-3 was predominantly found in the nucleus, whereas active caspase-7 and cleaved PARP localised mainly to perinuclear regions. It has been observed that in T cells active caspase-7 translocates to the endoplasmic reticulum upon apoptosis induction.29 Overall, these data indicate that caspase activation of lamina propria-infiltrating cells might play a role in IBD.

Caspase activation and poly(ADP-ribose)polymerase (PARP) cleavage are increased in tissue sections of patients with inflammatory bowel disease. Immunohistochemical detection of active caspase-3, active caspase-7 and caspase-cleaved PARP in representative colon tissues from a control person (A–C) and a patient with active Crohn's disease (D–F). Almost no immunostaining was obtained in control colon tissue, whereas lamina propria cells in the inflamed tissue of the patient with Crohn's disease showed intense staining with the antibodies (original magnification ×400).
Lamina propria Treg cells reveal increased apoptosis in patients with IBD
We next tried to characterise the cell type in the lamina propria undergoing apoptosis. By double staining for CD4/Foxp3 and CD25/Foxp3, we found a few scattered cells in control colon tissues that stained positively for the Treg markers (data not shown). Consistent with previous reports,15 17 30 patients with IBD revealed a considerably elevated number of Treg cells in the lamina propria as compared with control colon tissues. To investigate whether these lymphocytes undergo apoptosis, we performed a triple staining with TUNEL together with CD4 and Foxp3 antibodies. As expected, control biopsies (n=5) revealed almost no apoptosis in lamina propria cells (figure 2A–D). In contrast, we found increased apoptosis in lamina propria-infiltrating CD4+Foxp3+ cells in CD (n=5; figure 2E–H). Very similar results were obtained in colon tissue of patients with UC (n=5), whereas patients with microscopic colitis (n=4) revealed only low apoptosis of Treg cells (data not shown).
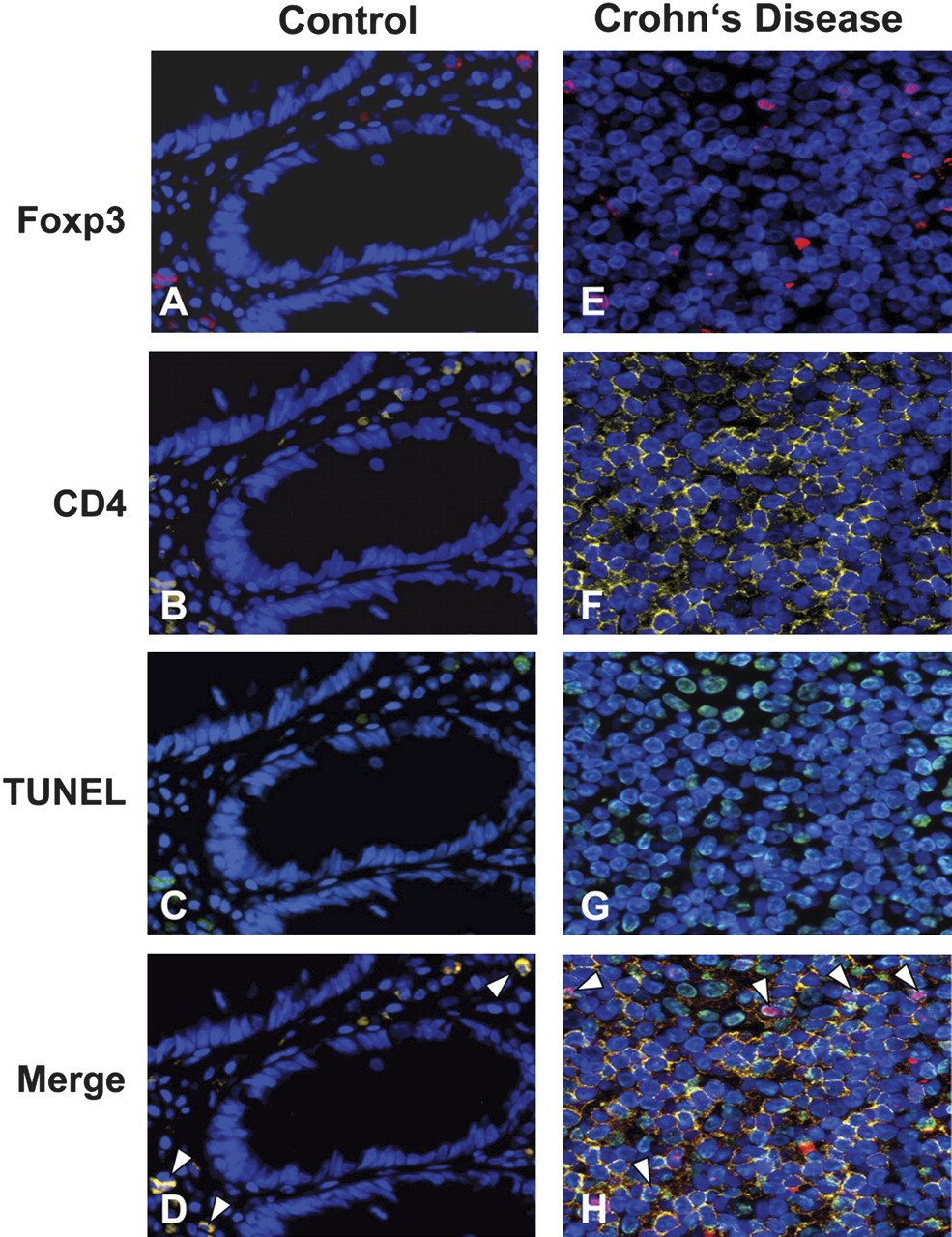
Elevated apoptosis of CD4+Foxp3+ cells in tissue sections of individuals with inflammatory bowel disease (IBD). Immunofluorescent triple staining of tissue sections from a representative control individual (A–D) and a patient with active Crohn's disease (E–H). Tissue sections were stained with 4′,6-diamidino-2-phenylindole (DAPI; blue), anti-Foxp3 (red; A, E), anti-CD4 (yellow; B, F) and TUNEL (terminal deoxynucleotidyl transferase dUTP nick end labelling) (green; C, G). A composite image (D, H) in which TUNEL was superimposed on CD4 and Foxp3 staining shows more triple-positive cells (arrowheads) in patients with IBD compared with control tissues. Magnification: ×400.

Quantification of CD4 T cell apoptosis in tissue sections of non-inflamed controls and patients with microscopic colitis (MC) or inflammatory bowel disease (IBD). The percentage of apoptosis of Foxp3-positive (A) and Foxp3-negative (B) CD4 T cells (mean±SEM) was assessed by counting four microscopic fields at ×400 magnification. Tissue sections from patients with ulcerative colitis (UC, n=5) or Crohn's disease (CD, n=5) showed significantly higher CD4 T cell apoptosis compared with non-inflamed controls (n=5) or patients with MC (n=4). **p<0.01, n.s., not significant.

Reduced frequency of CD4+CD25+Foxp3+ T cells in the peripheral blood of patients with inflammatory bowel disease (IBD). (A) Comparative assessment of the percentage of CD4+CD25highFoxp3+ T cells among the CD4 population in 10 IBD cases (Crohn's disease (CD) n=7, ulcerative colitis (UC) n=3) and 10 healthy controls. Lymphocytes were gated based on their scatter characteristics and stained for CD4, CD25 and Foxp3. The percentage of CD4+CD25highFoxp3+ regularoy T (Treg) cells among the CD4 population (mean±SEM; *p<0.05) is shown. (B) Representative FACS (fluorescence-activated cell sorting) plots of a control individual (left panel) and a patient with IBD (right panel). Numbers in each histogram indicate the percentage of CD25highFoxp3+ cells among the CD4+ T cells.

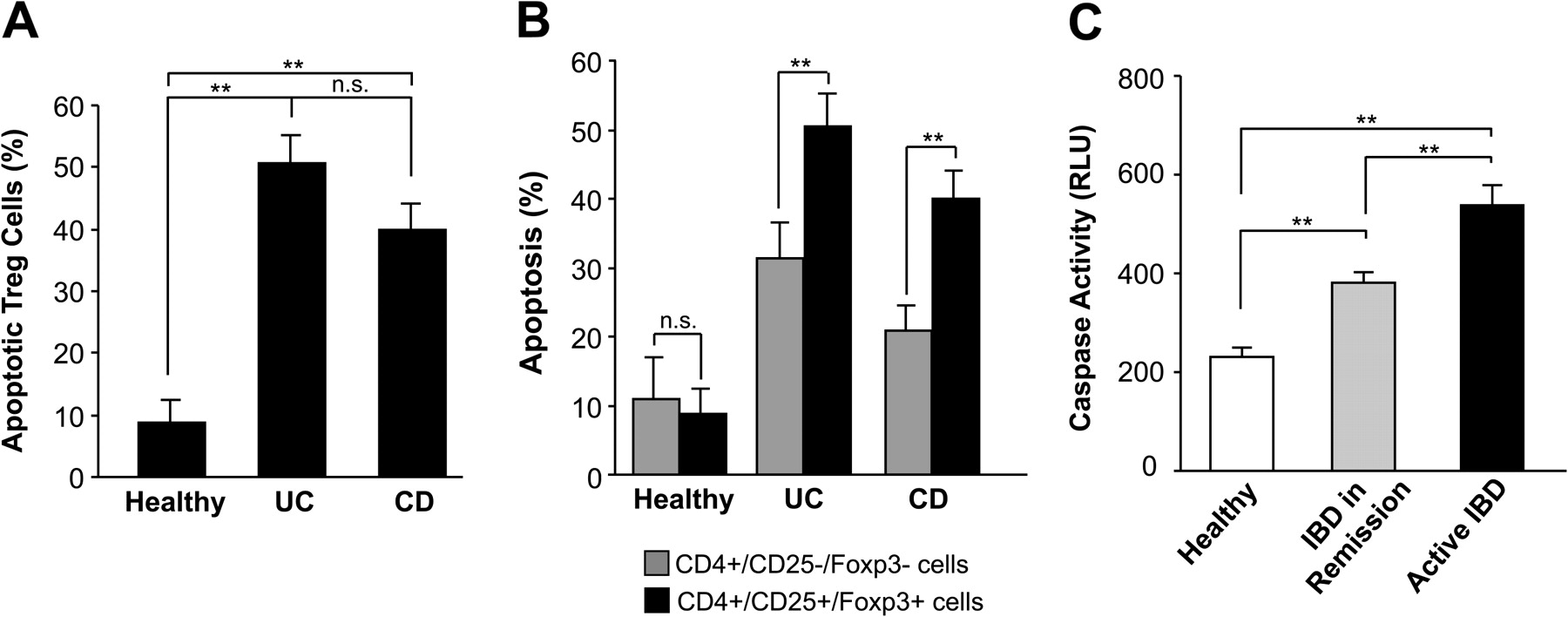
Elevated apoptosis of CD4+CD25+Foxp3+ regulatory T (Treg) cells in peripheral blood and enhanced serum caspase activity in patients with inflammatory bowel disease (IBD). (A) The percentage of apoptotic (annexin-V-positive) Treg cells in patients with ulcerative colitis (UC, n=24), Crohn's disease (CD, n=32) and healthy individuals (n=10) is shown. (B) Comparison of the percentage of apoptotic CD4+CD25+ T cells with the percentage of apoptotic CD4+CD25– T cells in healthy individuals (n=10), and patients with UC (n=24) and CD (n=32). (C) Luminometric detection of caspase activity in sera of patients with active IBD (n=66), inactive IBD (n=29) and controls (n=18). Patients with IBD showed higher serological caspase activity compared with healthy individuals. **p<0.01.

Inhibition of peripheral regulatory T (Treg) cell apoptosis during antitumour necrosis factor α (TNFα) treatment correlates with reduced serum caspase activity and disease activity. A representative example of a patient with Crohn's disease is shown. Apoptosis (annexin-V staining) of regularoy T (Treg) CD4+CD25highFoxp3+ cells (A), serum caspase activity (B), CRP levels (C) and CDAI (D) were analysed before (week 0) and during treatment with anti-TNFα (week 2 and 14). CRP, C-reactive protein; CDAI, Crohn's disease activity index; RLU, relative light units.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
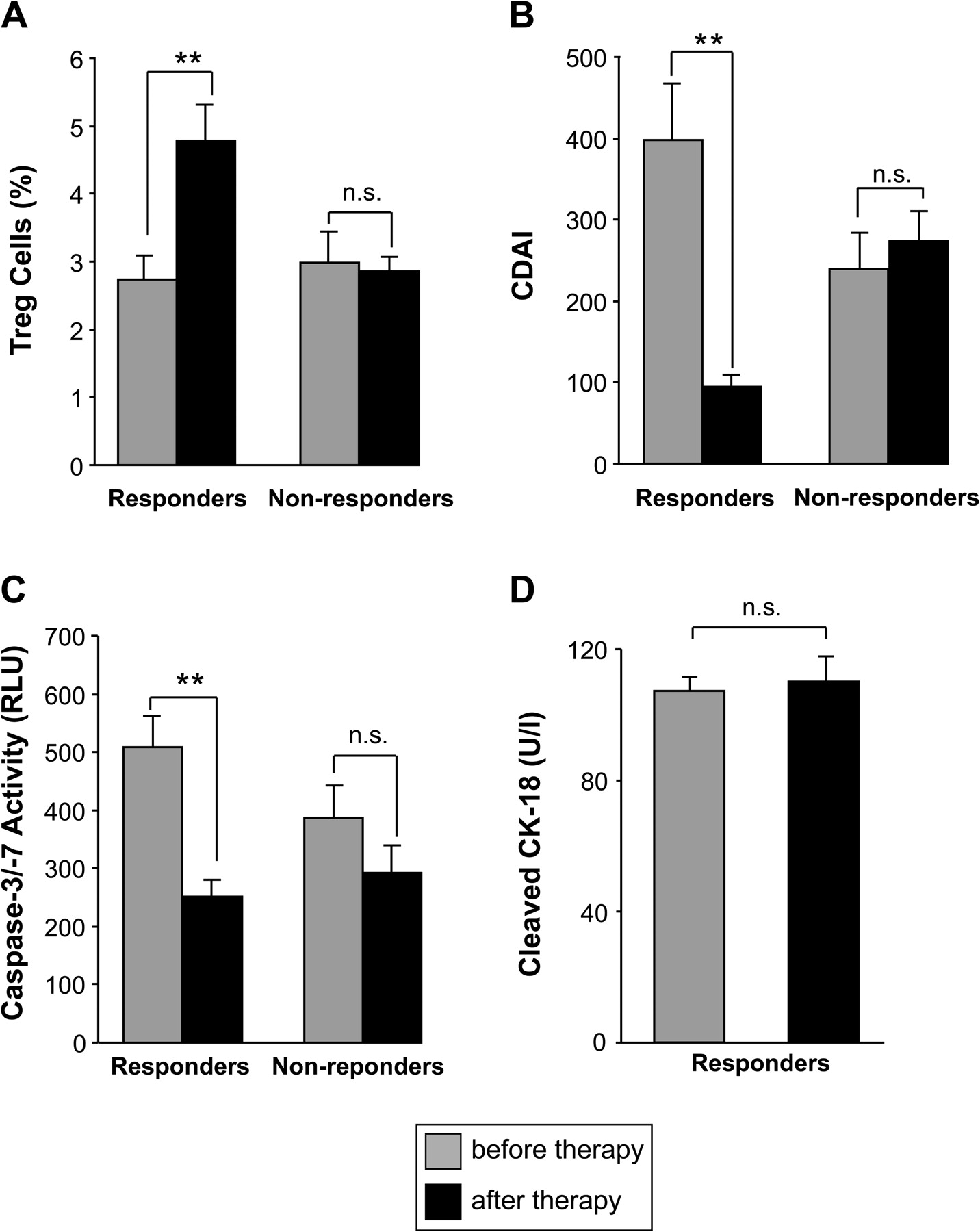
Restoration of CD4+CD25highFoxp3+ regulatory T (Treg) cell numbers and reduced serological caspase activity after successful antitumour necrosis factor α (TNFα) treatment. Flow cytometric determination of the percentage of peripheral blood Treg cells among CD4 cells (A), Crohn's disease activity index (CDAI) (B) and serological detection of caspase activity (C) is shown before and after anti-TNFα treatment in responding (n=6) and non-responding (n=5) patients with Crohn's disease. The frequency of Treg cells increased, whereas serological caspase activity declined after anti-TNFα-induced remission. Measurement of caspase-cleaved cytokeratin-18 did not reveal significant differences during anti-TNFα treatment (D), indicating that alterations of caspase activity were not caused by changes in epithelial cell viability. n.s., not significant; **p<0.01.
To quantify apoptotic CD4+Foxp3+ Treg cells in relation to the total number of Treg cells, CD4+Foxp3+ cells either with or without TUNEL reactivity were counted in the tissue sections (figure 3A). Non-inflamed control tissues (n=5) showed a mean of 3.5±2.2% apoptotic Treg cells compared with 17.9±2.0% in patients with microscopic colitis (n=4). Patients with IBD (CD n=5, UC n=5) revealed significantly (p<0.01) higher apoptosis rates of Treg cells, reaching 66.7±4.3% in CD and 62.0±5.1%, in UC, respectively. To determine whether the increased apoptosis in IBD was specific for Treg cells, we also determined the percentage of TUNEL reactivity in Foxp3-negative CD4 T cells. Figure 3B demonstrates a higher apoptosis of CD4+Foxp3– T cells in patients with CD (47.2±3.0%) and UC (48.1±11.7%) compared with microscopic colitis (11.3±2.3%) and non-inflamed controls (6.5±4.9%). These results indicate that although a considerable fraction of Foxp3-negative CD4 T cells underwent apoptosis, TUNEL reactivity was most prominent in the Foxp3-positive CD4 population. The difference in apoptosis between Foxp3-positive and negative CD4 T cells was significant (p<0.01) in patients with CD but not in those with UC.
Reduced frequency and increased apoptosis of Treg cells in the peripheral blood of patients with IBD
Having demonstrated increased caspase activation and apoptosis of Treg cells in the inflamed mucosa, we investigated alterations in the peripheral blood of patients with IBD. In control individuals (n=10), we found that on average 4.4±0.5% of the CD4+ T lymphocytes comprised Treg cells, as assessed by flow cytometry for CD4, CD25 and Foxp3 expression (figure 4). Interestingly and in line with a previous report,17 the percentage of peripheral Treg (CD4+CD25+Foxp3+) cells was significantly (p<0.05) reduced (2.7±0.4% of CD4+ T lymphocytes) in 10 analysed cases of active IBD compared with healthy individuals (figure 4A). Representative flow cytometric analyses of peripheral Treg cells in a control person and a patient with CD are shown in figure 4B. In addition to Foxp3 and high CD25 expression, Treg cells are characterised by a low or absent expression of CD127. A similar reduction of Treg cells was found in patients with IBD compared with healthy individuals, when cells were additionally stained for CD127 that was absent in >90% of the CD4+CD25+Foxp3+ population (data not shown).
The reduced frequency of peripheral Treg cells was proposed to reflect their enhanced outflow and recruitment into the mucosa.17 However, we found that a considerable fraction of the Treg cells revealed apoptosis, as detected by annexin-V staining (figure 5A). In line with a decreased Treg cell number (figure 4), apoptosis of peripheral blood Treg cells was significantly increased in patients with active UC (n=24) and CD (n=32) compared with control individuals (n=10). Representative fluorescence-activated cell sorting (FACS) analyses are shown in Supplementary figure 1. To analyse the ratio of regulatory to effector CD4 T cells we further compared the percentage of apoptosis in CD4+CD25– cells and Treg cells (figure 5B). Control individuals exhibited low apoptosis rates and no significant difference between apoptosis of CD4+CD25– cells (11.0±6.0%) and Treg cells (8.8±3.7%). In contrast, patients with UC showed significantly higher apoptosis of Treg cells (50.5±4.8%) compared with effector CD4 T cells (31.4±5.2%). Similar results were observed in patients with CD (40.0±4.1% vs 20.8±3.8%). These data indicate a disturbed T cell balance due to a significantly higher apoptosis rate of Treg cells compared with effector CD4 T cells.
Since increased apoptosis might be associated with the release of active caspases into serum, we also determined caspase activity in sera of patients with IBD using a previously established assay.26 To this end, serum samples were incubated with the substrate DEVD-aminoluciferin which upon caspase cleavage generates a luminescent signal (figure 5C). Patients with IBD indeed revealed a strongly increased caspase activity in the serum. Patients with active IBD (n=66) showed significantly higher caspase activity (540±39 relative light units (RLU)) compared with patients with IBD in remission (n=29; 380±21 RLU) or control individuals (n=18; 231±17 RLU). No significant differences in caspase activity were observed between CD and UC, either in patients in remission or in those with active disease (data not shown). The elevated caspase activity in patients with IBD might be due to increased apoptosis of Treg cells but, given their low percentage, presumably also of other T cells. Although the DEVD substrate is preferentially cleaved by apoptotic caspase-3 and -7, we can also not exclude that inflammatory caspases (eg, caspase-1) might contribute to the increased serum activities in patients with IBD.
Peripheral Treg cell apoptosis and serological caspase activity in the course of anti-TNFα treatment
In autoimmune diseases such as rheumatoid arthritis, it was proposed that TNFα inhibition might eventually restore the number of peripheral Treg cells or promote their differentiation.31 We therefore investigated the Treg cell apoptosis and caspase activation in blood samples of patients with CD during anti-TNFα treatment. Anti-TNFα antibody was administered once at weeks 0, 2 and 6 and then every 8 weeks. Blood samples were taken immediately before and 2 and 14 weeks after initiation of anti-TNFα treatment. Although the data varied between individual patients, we consistently found reduced apoptosis in peripheral CD4+CD25highFoxp3+ (Treg) cells together with declined caspase activity upon anti-TNFα treatment. A representative example of a patient responding to anti-TNFα is shown in figure 6. During anti-TNFα treatment the apoptosis rate of Treg cells decreased from 25.6% before treatment to 12.6% after 14 weeks of anti-TNFα treatment (figure 6A). The decreased apoptosis of Treg cells was accompanied by reduced serum caspase activity (figure 6B) as well as by diminished levels of C-reactive protein (CRP) (figure 6C) and CDAI (figure 6D) that were determined as clinical markers of disease activity. These data indicate that the treatment response—ie, resolution of inflammatory disease activity—is associated with decreased apoptosis of Treg cells and serum caspase activity.
To substantiate further the finding of decreased Treg cell apoptosis in patients responding to anti-TNFα, we investigated the number of Treg cells before and during anti-TNFα treatment in patients with CD, either responding (n=6) or not responding (n=5) to anti-TNFα. These analyses verified our previous finding and revealed a significant (p<0.01) increase of the Treg cell frequency (4.8±0.5% Treg cells among CD4 cells) after anti-TNFα treatment compared with the low percentage of Treg cells before treatment (2.7±0.4% of Treg cells) in anti-TNFα responders (figure 7A). In contrast, in non-responding patients the frequency of peripheral Treg cells remained largely unchanged (2.9±0.4% before and 2.8±0.2% after anti-TNFα treatment). In line with this, responders showed a significant (p<0.05) decrease of Treg cell apoptosis (9.8±1.9%) after successful treatment compared with the high apoptosis rate (25.5±0.4%) before anti-TNFα application, whereas Treg cell apoptosis in non-responding patients remained largely unchanged (data not shown). Moreover, in responding patients the decreased apoptosis was confined to Treg cells, but not observed in the effector T cells (data not shown). The increase of Treg cells in the responders was associated with a normalisation of clinical parameters of disease activity, for example CDAI (before treatment 398±70, after treatment 94±16; figure 7B) and CRP levels (data not shown). In line with, caspase activity in serum of responding patients was also significantly (p<0.01) decreased (252±28 RLU) after anti-TNFα-induced remission compared with values before treatment (509±54 RLU; figure 7C). Non-responders, in contrast, showed no significant decrease of caspase activity in the course of treatment (386±56 RLU vs 292±46 RLU, figure 7C). In addition, we determined the serum levels of caspase-cleaved cytokeratin-18 (M30 antigen), which is often used as a serum marker for apoptosis of epithelial cells.28 Whereas caspase activity was enhanced in patients with IBD and reduced after anti-TNFα treatment, we found normal M30 levels that were also not affected by anti-TNFα, indicating that the increased caspase activity was not caused by tissue damage of epithelial cells (figure 7D). Altogether, these results suggest that increased apoptosis of Treg cells plays a potentially important role in IBD pathogenesis and can be reversed by anti-TNFα treatment.
Discussion
The pathogenesis of IBD is complex and involves defects in the mucosal barrier function and immune system. Despite many important findings in recent years, it is still largely unknown why the mucosal immune response is over-reactive in IBD, and how current medical treatment (ie, with anti-TNFα antibodies) affects the mucosal immune system. In this study, we investigated apoptosis of CD4+CD25highFoxp3+ cells (referred to as Treg cells) as a possibly contributing factor to the dysregulated immune response in patients with IBD. While we could demonstrate a lack of caspase activation in the normal mucosa, lamina propria cells in patients with active CD showed an intense staining for activated caspases and caspase-cleaved PARP. Further characterisation revealed CD4+Foxp3+ Treg cells as a prominent apoptotic cell type in the gut. Duchmann and co-workers17 as well as other groups15 30 previously reported an accumulation of Treg cells in the lamina propria, which was interestingly significantly lower in IBD than in other intestinal inflammatory diseases including diverticulitis. In our study, we found higher apoptosis of CD4+Foxp3+ (Treg) cells in tissue samples of patients with active IBD compared with patients with diverticulitis. Our results therefore suggest that counter-regulation of inflammation by Treg cells in IBD might be relatively ineffective because of their increased rate of apoptosis.
Reduced peripheral Treg cell numbers have been found associated with both active CD and UC, and shown to correlate inversely with disease activity.17 32 Consistent with these reports, we also found a significantly decreased frequency of peripheral CD4+CD25highFoxp3+ cells in patients with IBD compared with healthy individuals. The decreased number of circulating Treg cells was thought to reflect their accumulation in the intestine. However, we demonstrate for the first time a significantly higher rate of apoptosis in intestinal and peripheral blood Treg cells of patients with IBD. Moreover, we demonstrate an increased caspase activity in the serum of patients with IBD. This caspase activity might be due to increased T cell apoptosis, although non-apoptotic caspases that are activated during inflammation might also have contributed to the increased values. Interestingly, whereas caspase activity was enhanced in IBD sera, we did not find elevated levels of caspase-cleaved cytokeratin-18, a serum marker for apoptosis of epithelial cells. The data suggest that peripheral Treg cells decrease not only due to their outflow into the mucosa, but also owing to their enhanced rate of apoptosis.
In contrast to IBD, no reduction in numbers of Treg cells has been consistently found in patients with rheumatoid arthritis. There is only a single report showing a reduced number of CD4+CD25+ cells in rheumatoid arthritis as compared with healthy individuals.33 Most studies suggest that a functional impairment rather than a numerical defect is involved in the dysregulated immune response of rheumatoid arthritis. Although we have not investigated the suppressor activity and have defined Treg cells solely by their CD4/CD25/Foxp3 profile, several previous studies demonstrated that peripheral blood and mucosal Treg cells are functionally normal in IBD.15 30 It was also reported that CD4+CD25+ lamina propria T cells in IBD have normal regulatory activities, since they suppress proliferation and cytokine production of CD4+CD25− cells.34
There are several possibilities that could be involved in the increased apoptosis susceptibility of Treg cells in IBD. Data from experimental colitis models revealed that inflammation-induced depletion of Foxp3+ Treg cells occurs via CD95-mediated apoptosis.35 Whether patients with IBD show increased CD95-mediated apoptosis of their Treg cells remains to be elucidated. Another explanation for the insufficient Treg cell supply might be that colitis-driving effector T cells negatively influence Treg cell development. Evidence for this hypothesis comes from T cell-deficient Tgε26 mice that are characterised by a rudimentary thymus.36 37 Although bone marrow repopulation can restore most thymic functions, those mice still suffer from a numeric defect of Treg cells, resulting in chronic colitis. Interestingly, the onset of colitis in this model is accompanied by a diminished thymic cellularity, whereas treatment with anti-TNFα or Treg cells prevents thymic collapse and leads to development of functional Treg cells.38 39 These results implicate a colitis-induced feedback mechanism leading to a defective thymic production of Treg cells in genetically susceptible hosts.
In the present study we demonstrate for the first time that in patients with CD responding to anti-TNFα treatment the apoptosis rate of peripheral blood CD4+CD25highFoxp3+ (Treg) cells as well as serological caspase activity are diminished and accompanied by decreased CRP and CDAI levels. These data indicate that treatment response (ie, resolution of inflammatory disease activity) is associated with decreased apoptosis of Treg cells. In line with this is our finding that the reversal of spontaneous apoptosis by anti-TNFα correlates with a significant increase in the number of peripheral CD4+CD25highFoxp3+ (Treg) cells, which was not observed in patients with treatment failure with anti-TNFα. Similar to peripheral blood, we also found a decreased apoptosis and increased number of CD4+Foxp3+ cells in the mucosa of patients with CD after successful anti-TNFα treatment (Supplementary figure 2). These results imply that the increase in Treg cell number might be responsible for the disease resolution.
TNFα is a pleiotropic cytokine critical for inflammation and host defence against various pathogens. Thus, the beneficial effects of TNFα blockade might be mediated by several mechanisms including inhibiting the production of proinflammatory mediators and CD4+ T cell effector function. There are indications that in certain autoimmune diseases TNFα inhibition affects Treg cell differentiation. In rheumatoid arthritis, anti-TNFα treatment might directly restore the compromised suppressor function of Treg cells.39 Another study suggested that TNFα antibodies could not restore the defective function of natural Treg cells, but promoted T cell differentiation to a new population of Treg cells in the peripheral blood.22
In IBD the restored Treg cell number and decline of clinical disease activity strongly support the assumption that intestinal Treg cell apoptosis might be decreased by TNFα inhibition and therefore constitute an additional mechanism for the beneficial effects of anti-TNFα treatment. This assumption is supported by an in vitro study showing that TNFα blockade augments the percentage of Treg cells.40 TNFα might directly induce apoptosis of Treg cells by binding to TNF receptor-2 that is expressed on Tregs and upregulated by TNFα.22 23 In addition, proinflammatory cytokines and impaired expression of IL-10 or transforming growth factor β (TGFβ) can abrogate the suppressive activity of Treg cells.41 In addition to neutralising TNFα, anti-TNFα treatment can suppress the production of IL-6 and IL-1β that are also implicated in rendering Treg cells defective. Anti-TNFα antibodies have further been reported to restore the function of Tregs, to increase Foxp3 expression and to decrease TNF receptor-2 expression.22 39 A very recent report demonstrated that anti-TNFα treatment of IBD, similar to what was found in our study, restored the number of peripheral Treg cells but, surprisingly, caused a diminished mucosal expression of Foxp3, indicative of a decreased Treg cell number in the mucosa.42 Unfortunately, the authors did not investigate T cell apoptosis, and, moreover, employed different antibodies for measurements of either peripheral or mucosal Foxp3 expression, which might explain the differences from our study. Certainly, the cellular pathways and signalling mechanisms that might be directly or indirectly affected by TNFα inhibition require further investigation.
In summary, our data suggest that increased apoptosis of peripheral and mucosal Treg cells is a potentially important contributing factor in IBD pathogenesis. Anti-TNFα treatment resulted in decreased apoptosis of CD4+CD25highFoxp3+ (Treg) cells with subsequent enhancement of Treg cell number. Since there is neither a consensus about the duration of anti-TNFα treatment nor a reliable disease activity marker, detection of Treg cell apoptosis might allow for monitoring the disease activity and treatment response to anti-TNFα therapy.
References
Supplementary materials
Online only appeddix
Files in this Data Supplement:
Footnotes
CV and MA contributed equally to this work.
Funding This study was supported by the Deutsche Forschungsgemeinschaft (SFB773, SFB685, TRR77, GRK1302, BA 2092/2-1, BA 2092/9-1) and the Federal Ministry of Education and Research (BMBF, No. 01FP09104B and AID-Net project).
Competing interests None.
Ethics approval This study was conducted with the approval of the Hannover Medical School and University of Heidelberg.
Provenance and peer review Not commissioned; externally peer reviewed.