Article Text

Abstract
Our life expectancy is increasing, leading to a rise in the ageing population. Ageing is associated with a decline in physiological function and adaptive capacity. Altered GI physiology can affect the amount and types of nutrients digested and absorbed as well as impact the intestinal microbiota. The intestinal microbiota is considered a key player in our health, and a variety of studies have reported that microbiota composition is changing during ageing. Since ageing is associated with a decline in GI function and adaptive capacity, it is crucial to obtain insights into this decline and how this is related to the intestinal microbiota in the elderly. Hence, in this review we focus on age-related changes in GI physiology and function, changes of the intestinal microbiota with ageing and frailty, how these are associated and how intestinal microbiota-targeted interventions may counteract these changes.
- ageing
- gastrointestinal physiology
- intestinal bacteria
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Ageing is associated with alterations in GI function, on both the organ and cellular level, including an impaired immune function.
The composition of the intestinal microbiota changes during ageing.
What are the new findings?
The direct impact of age-related alterations in GI function on nutrient digestion and absorption, as well as on the intestinal microbiota composition and functionality, is largely unknown.
Intestinal microbiota perturbations in the elderly are more likely to be associated with health status, medication use and lifestyle factors, rather than with chronological ageing per se.
How might it impact on clinical practice in the foreseeable future?
The intestinal microbiota could be a promising target to improve health outcome in relevant subgroups of elderly.
More studies are needed on the impact of manipulating the intestinal microbiota, using a targeted approach and clear focus on well-characterised and relevant subgroups of elderly (such as frail elderly, specific comorbid conditions and/or medication use).
Introduction
Although geographical differences exist, the overall life expectancy is increasing worldwide, leading to a steady rise in the ageing population. In Europe, the proportion of individuals aged 65 years and over has been estimated to increase from 17% in 2010 to 30% in 2060.1 Ageing is associated with an inevitable time-dependent decline in physiological function and adaptive capacity, as a result of lifelong accumulating molecular and cellular damages.2–4 Several studies have shown that this decline is host-specific and can be influenced by various factors such as host genetics, lifestyle (eg, diet and smoking), sociodemographics (eg, age, socioeconomic status and ethnic background), living situation (eg, community-dwelling or institutionalised)5 and comorbidities (including medication use), contributing to large heterogeneity in the elderly population. Whereas conventionally elderly were defined as being >65 years of age, nowadays the overall rate of biological ageing is decreasing, and cut-offs as well as age definitions vary widely. In scientific research and in clinical settings, it is therefore relevant to assess physiological functionality rather than focusing on chronological age. Some studies include subjects with specific comorbidities or (pre-)frail elderly to address differences in age-related physiology. Others focus on centenarians to get further insight into genetic and lifestyle factors associated with longevity and resilience to disease.6
Frailty is ’a syndrome of decreased reserve and resistance to stressors, resulting from cumulative declines across multiple physiological systems, causing vulnerability to adverse outcomes’ as defined by Fried et al.7 Applying this definition in a meta-analysis of 2009, the pooled prevalence of frail elderly in community-dwelling adults aged 65 years and older in Europe was 17.0%.8 The frailty phenotype of Fried et al 7 has been operationalised to assess physical frailty by evaluating unintentional weight loss, self-reported exhaustion, weakness (eg, by handgrip strength), slow walking speed and low physical activity. Besides these widely adopted Fried criteria, several multidisciplinary scores exist that include measures of medical, psychological, cognitive, functional and/or social loss.9 In an extensive systematic review, 79 different frailty assessment tools have been identified, although a ‘gold’ standard is lacking.9 Since frailty is associated with an increased risk of negative health outcomes leading to disability and impaired quality of life,10 the increasing group of frail elderly leads to more direct (eg, consultations, diagnostic procedures, hospitalisations, medication use) and indirect healthcare costs (eg, social and daily support). Therefore, further insights into modifiable factors and preventive strategies are highly relevant.
The intestinal microbiota, which is the collection of microbes found in the intestine, may be a promising target as it clearly links to a myriad of host functions, is affected by environmental factors, and perturbations have been reported in the ageing population. Nutritional interventions, aiming to modulate intestinal microbiota composition and functionality, may contribute to intestinal health and general well-being (of the ageing population). Therefore, our aim was to review (1) the current knowledge on the effect of ageing on GI physiology and on intestinal microbiota, (2) whether other factors besides ageing, such as frailty, affect GI physiology and intestinal microbiota and (3) potential targets to counteract the changes in GI physiology and intestinal microbiota observed in elderly and/or frailty. Hence, current knowledge on GI physiology and function, intestinal microbiota composition and activity and manipulation of the intestinal microbiota in relation to ageing and frailty will be summarised, paying specific attention to age-definitions and associated health status.
Physiology and function of the ageing GI tract
The GI tract has a core function for the human body as it ensures adequate digestion and absorption of nutrients, involving a sequence of events from the mouth until the anus. These are facilitated by GI motility and digestive secretions, and regulated by neural and hormonal control. Age-related functional declines have been reported for some of the organs involved (figure 1).

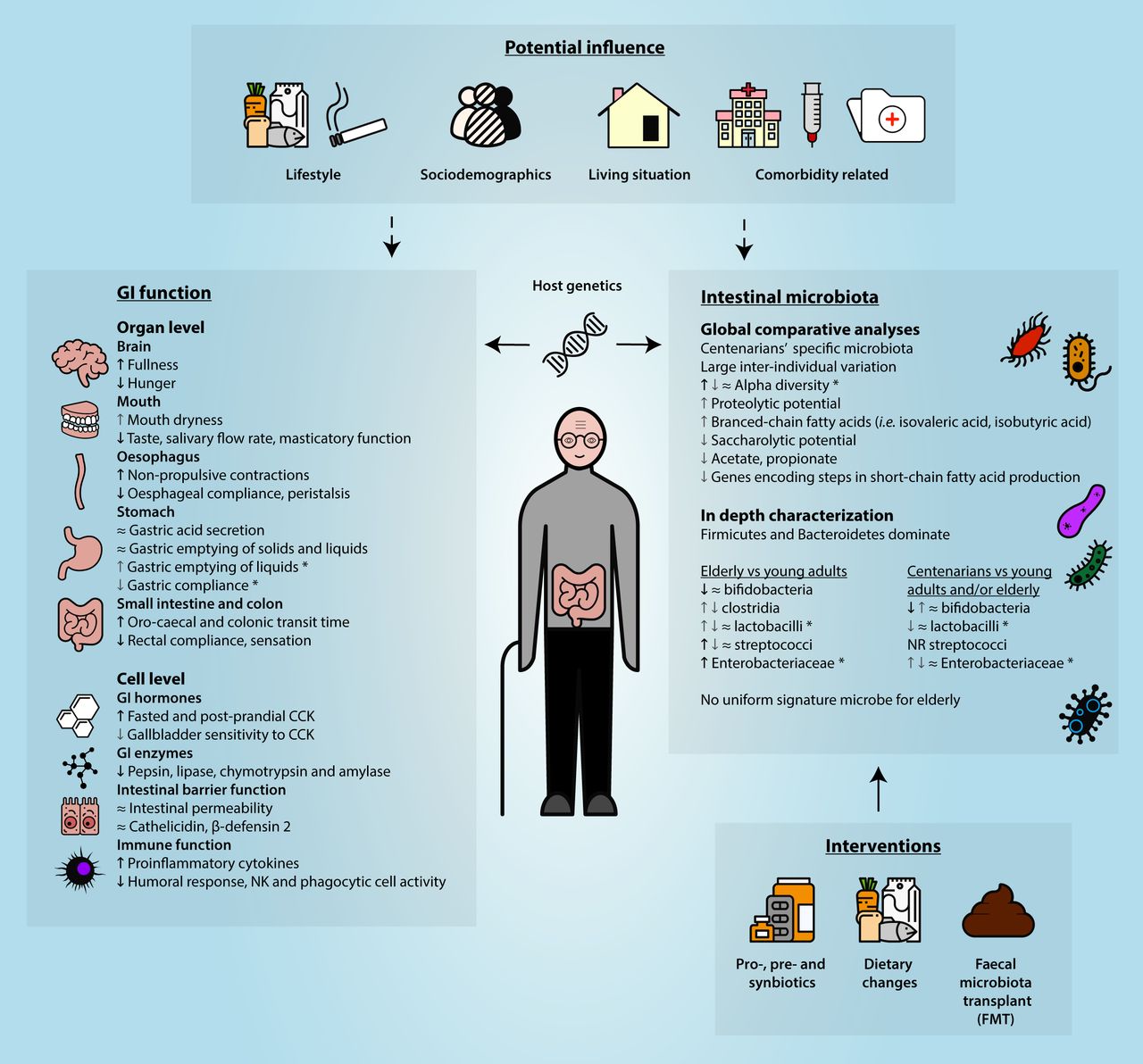{kind=link}
Non-exclusive listing of key changes in GI function and the intestinal microbiota during ageing, including potential influencing factors as well as interventions to beneficially manipulate the intestinal microbiota. *Frailty related; ≈no significant difference;  a few or the minority of studies showing a significant increase;
a few or the minority of studies showing a significant increase;  several or the majority of studies showing a significant increase;
several or the majority of studies showing a significant increase;  a few or the minority of studies showing a significant decrease;
a few or the minority of studies showing a significant decrease;  several or the majority of studies showing a significant decrease. CCK, cholecystokinin; NK, natural killer; NR, not reported.
several or the majority of studies showing a significant decrease. CCK, cholecystokinin; NK, natural killer; NR, not reported.
In the mouth, masticatory function and taste are often found to be impaired in elderly in general.11 Moreover, Watanabe et al 12 found that frail elderly had significantly fewer teeth, lower occlusal force and muscle thickness when compared with healthy and prefrail elderly, indicating that impaired masticatory function becomes more pronounced with frailty.12 Swallowing problems were reported by 10%–30%, in a heterogeneous group of individuals aged 65 years and over.13 Swallowing depends among others on the presence of saliva. Results from a meta-analysis, including 47 original controlled studies, showed that salivary flow rate decreases with healthy ageing, resulting from degenerative changes of cellular structures of the salivary glands.14 While reduced salivary secretion has been associated with medication use such as anticholinergic drugs, psychotropic drugs, antihistamines and diuretics, subanalyses showed that medication use does not impact salivary flow rate.14 Frailty criteria were, however, not taken into account. A recent study by Rogus-Pulia et al 15 showed that perception of mouth dryness was also associated with increasing age, although potential contributing factors like comorbidities and medication use were not addressed.15 The observed dryness is probably due to compositional changes in saliva, which may negatively impact thickness and adherence of the oral salivary film.15 16 Increased impairment in mouth functions as reported for elderly in general, can contribute to food avoidance, reduced or altered intake (eg, more easily digestible food) and eventually poor nutritional status.11 This can also impact intestinal microbiota composition and activity. Deteriorated oesophageal functions repeatedly reported in elderly in general, include reduced peristalsis,17 18 increased number of non-propulsive contractions18 and decreased compliance of the oesophagus.18 These alterations can be found already from the age of 40 years.19 Healthy ageing was not or only to a modest extent found to be associated with slowing of gastric emptying of both solids and liquids.20 21 In frail compared with non-frail elderly, gastric emptying of liquids was even found to be enhanced.21 Furthermore, the gastric compliance was reduced, while gallbladder emptying and oro-caecal transit time were not different between frail and non-frail elderly.21 On the other hand, in a recent review it was stated that healthy elderly have a longer oro-caecal and colonic transit time than healthy young adults.22 The decreased rectal compliance and decreased rectal sensation associates with healthy ageing, and may contribute to complaints such as constipation.18 Several studies have provided evidence for alterations in motility associated with healthy ageing, being most pronounced for the oesophagus and large intestine. It should be acknowledged that increased intestinal transit time affects composition and activity of intestinal microbiota, both in vitro23 and in vivo24, but data in elderly or frailty are lacking.
Most GI functions, including the secretion of gastric juice, bile and digestive enzymes, as well as GI motility, are regulated by a complex interplay of hormonal and neuronal factors, involving the central and the enteric nervous system. In the stomach, marked alterations in gastric acid secretion could not be demonstrated in healthy elderly.25 Although, the higher prevalence of atrophic gastritis and proton pump inhibitor use in elderly20 will lead to decreased acid production and subsequently bacterial overgrowth in subgroups. Rémond et al 26 reviewed studies on the effects of ageing on excretion of digestive secretions and found that bicarbonate secretion, as well as enzyme concentrations of pepsin in the stomach, and lipase, chymotrypsin and amylase in duodenal fluids were lower in healthy elderly compared with young adults. However, bile acid secretion was not affected by ageing.26 In an extensive review on (an)orexigenic GI hormones, strongest evidence was found for elevated fasted and postprandial concentrations of cholecystokinin (CCK) in elderly in general compared with younger adults.27 Although gallbladder emptying was similar, gallbladder sensitivity to endogenous CCK was significantly reduced in healthy elderly compared with young adults.28 Evidence on the effects of ageing on peptide YY, glucagon-like peptide-1 and ghrelin levels were inconclusive, partly as a consequence of different methodologies used, whereas pancreatic polypeptide and oxyntomodulin are hardly studied.27 Data on other relevant GI hormones and peptides (eg, secretin, gastrin, motilin, somatostatin, chromogranin A) as well as specifically addressing subgroups of elderly are scarce. More studies using standardised methods and clear definitions of the target population are needed. The enteric nervous system also plays an important role in the regulation of motility, secretions and local blood flood, by bidirectional communication between the brain and the gut and by local reflexes. Several rodent studies found the ageing process to induce neuronal loss as determined by lower neuronal density in oesophageal, small intestinal and colonic tissue.20 This effect was more pronounced in cholinergic when compared with nitrergic myenteric neurons.29 Human data are, however, lacking.
Although several of the above processes will impact GI function, studies on intestinal absorption of nutrients per se are largely lacking. In rodent studies, ageing was associated with a decrease in surface area because of villus degeneration20 and malabsorption of carbohydrates, lipids, proteins, minerals and vitamins,30 but these findings need confirmation in humans. Food intake was, however, found to be altered in healthy elderly, which was associated with decreased hunger and appetite scores.27 A meta-analysis including 59 studies confirmed that hunger scores were 25% and 39% lower after overnight fasting and in postprandial state, respectively, and fullness 37% higher in healthy elderly compared with young adults.31 Furthermore, reduced nutritional status was found to be related to frailty in elderly.32
Apart from the segment-specific functions with regard to digestion and absorption, an adequate GI barrier function is pivotal for protection against the external environment, including epithelial integrity, mucus and defensin secretion, as well as the gut-associated lymphoid tissue. Although studies in primates suggest an impaired epithelial barrier function in older animals,33 Wilms et al 34 did not observe differences between healthy elderly versus adults based on results from a combined in vivo (using the multisugar permeability test) and ex vivo approach (evaluating colonic biopsies in Ussing chambers).34 Although not specifically studied in elderly, factors associated with ageing such as non-steroidal anti-inflammatory drugs (NSAID) use, alcohol intake, obesity and diabetes have, however, been shown to increase intestinal permeability. The ‘chemical’ barrier is hardly studied in elderly. One study showed that serum levels of the host defensive peptides cathelicidin and β-defensin 2 were similar between healthy elderly and young adults.35 Data on mucosal and/or faecal samples as well as on the role of other antimicrobial peptides or mucus and analyses in frail elderly specifically are lacking. Many studies explored the impact of ageing on the intestinal immune response. It has repeatedly been shown that the human intestinal mucosal immune system is compromised with ageing, as nicely reviewed by Mabbott et al.36 Alterations in dendritic cell subsets have been reported in elderly in general.36 Furthermore, healthy ageing is characterised among others by a pro-inflammatory cytokine profile (‘inflammaging’, ie, increased levels of among others tumour necrosis factor-α and interleukin-6), a decreased humoral response such as reduced secretory IgA levels and decreased natural killing and phagocytic cell activity.37 This will be further affected by immune-related comorbid conditions and/or frailty.37 The observed immunosenescence can contribute to the increased risk of recurrent and persistent infections reported in the elderly.37
Intestinal microbiota in elderly
Ageing is associated with several alterations in GI physiology and function, which can impact the amount and types of nutrients delivered to the small intestine and colon, thereby affecting intestinal microbiota composition and functionality in these segments. Additionally, the altered immune function will impact host-microbe interactions, which can also contribute to alterations in intestinal microbiota composition and functionality in this population.
The GI tract encompasses different biotic environments. Although different types of microorganisms, such as archaea and fungi, may play a role in intestinal health,38 most studies focus on bacteria. Furthermore, it is evident that different locations of the GI tract harbour distinct microbial communities,39 but determination of the bacterial composition is mostly performed on faecal samples as their collection is non-invasive and feasible for large ((pre)clinical) populations. Nevertheless, other intestinal sites can be sampled using luminal brushes, rectal swabs, colonic lavage and mucosal biopsies as reviewed earlier.38 In elderly, studies have mainly focused on analyses of the microbiota composition in faecal samples, which is generally considered to be representative for the distal large intestinal content. In this section, we summarise the current knowledge on microbiota composition of elderly, and discuss whether the faecal microbiota of young adults is different from elderly, paying special attention to age-related health status (ie, (pre)frail and centenarians) and confounding factors.
Global faecal microbiota comparative analyses
A wide variety of studies have compared the faecal microbiota composition in elderly versus young adults and centenarians. In online supplementary table S1, a non-exclusive overview of studies investigating the faecal microbiota profile of elderly is given. Their overall microbiota composition was generally visualised and verified by ordination and multivariate analysis, for example, significant age group-based separation in diagrams. At phylum level, the faecal microbiota of young adults and elderly was found to be rather similar in some studies,40–43 whereas others reported significant differences.40 44 45 Remarkably, the faecal microbiota of centenarians, who are reported to have a lower incidence46 of chronic illnesses than those elderly aged 80–99 years and considered a ‘successful’ ageing model,47 was reported to be different from that of non-institutionalised elderly aged 70 years.42 Although these studies were conducted in different countries, a clear link with the geographic origin cannot be observed. Part of the contradictory findings between young adults and elderly could be due to differences in recruitment strategy (in/exclusion criteria), age definitions and confounding factors, such as comorbidity, medication use, lifestyle and socioeconomic factors, hampering an adequate comparison. For instance, the microbiota composition of smokers was different from that of non-smokers, aged 20–59 years.48 The effect of smoking in combination with ageing has not been reported to date. Overall, the microbiota of elderly is highly variable.45 Therefore, it is hard to define a typical microbiota of elderly and that of centenarians.
Supplemental material
Observations with regard to the effect of ageing on the alpha diversity, which is compositional complexity based on richness and evenness of the microbial ecosystem, vary, especially due to frailty and within the group of centenarians (table 1). Initially, alpha diversity was found to decline during ageing,49 50 which was mainly based on cultivation and classical 16S ribosomal RNA (rRNA) gene approaches. However, this observation could not be confirmed by high-throughput 16S rRNA gene sequencing and phylogenetic microarrays. Several studies even reported higher alpha diversity51–54 in the microbiota of community dwelling elderly versus young adults, while others reported no significant differences.42 55 56 A high alpha diversity has often been suggested to be associated with better homeostasis and resilience to disturbance.57 In terms of centenarians, their alpha diversity was reported to be higher than that of elderly,43 58 59 but not exclusively.42 51 59 Moreover, a broad range of confounding factors can affect the varying microbiota alpha diversity observed, including host and/or lifestyle factors. For instance, a decreased alpha diversity was reported for smokers compared with non-smokers.48 In addition, although not exclusively,60 61 lower alpha diversity has been reported to be associated with increased53 55 56 frailty (table 2), which suggests that the health status of elderly rather than ageing itself is associated with a lower alpha diversity of the faecal microbiota.
Ageing-associated changes in alpha diversity of faecal microbiota in elderly and centenarians
Summary of studies associating faecal microbiota profiles with frailty
In-depth characterisation of faecal microbiota composition
The first paper that associated the dynamics of several bacterial genera during life was published by Mitsuoka62 in 1990. This culture-based hallmark paper described that in comparison to young adults, the faecal microbiota of elderly comprised a lower abundance of bifidobacteria, whereas clostridia, lactobacilli, streptococci, Enterobacteriaceae were increased. In this review, we compared and contrasted subsequent culture-based as well as culture-independent studies with respect to these identified ageing-associated bacterial groups.
The reduction in the abundance of bifidobacteria in the faecal microbiota of elderly has been confirmed in many studies,40 43 49 63–65 irrespective of elderly or frailty definitions, although not exclusively.42 43 66 Surprisingly, a decreased abundance of bifidobacteria has also been reported in centenarians (ie, 99–104 years), compared with young adults65 or elderly (>65 years),43 whereas an increased abundance (of bifidobacteria) was reported in (super)centenarians (>105 years).43 In an Italian cohort, no significant differences in bifidobacteria abundance were observed between elderly and centenarians (ie, 99–104 years).42 The abundance of bifidobacteria was reported to be lower in the subgroup of institutionalised geriatric elderly,49 in the subgroup of hospitalised elderly64 and in the subgroup of Clostridium difficile-associated disease (CDAD) elderly,63 compared with healthy elderly and young adults. Furthermore, the abundance of bifidobacteria was higher in another Italian cohort than in Swedish, German and French cohorts, irrespective of age (ie, both young adults and elderly), which was concluded to be due to differences in dietary habits.40 These observations indicate that comorbidity and habitual diet may affect the alterations in the abundance of bifidobacteria during ageing, but to what extent and how they could contribute to the observed changes is still not clear.
Clostridia, lactobacilli and streptococci belong to the Firmicutes phylum (often referred to as Gram-positive bacteria with low guanine-cytosine content in their DNA). In line with observations in young adults, the phyla Firmicutes and Bacteroidetes together form the most dominant fraction of the microbiota in elderly. However, whether Firmicutes51 67 68 or Bacteroidetes44 45 59 is the most dominant phylum, differs between studies.
The number of studies that has reported an effect of ageing on the abundance of clostridia is rather small when compared with other bacterial groups. This can in part be attributed to the continuous reclassification and renaming of anaerobic spore-forming bacterial isolates after the introduction of 16S rRNA gene-based taxonomy that were traditionally named Clostridium. Although increases and reductions in the abundance of clostridia have been described in the faecal microbiota of elderly versus young adults,64 69 comparative analyses between studies is hampered by this continuous reclassification.
The reported increase in abundance of lactobacilli during ageing by Mitsuoka et al could not be confirmed in other culture-based studies,63 64 70 whereas it was confirmed in several culture-independent studies.40 69 71 The abundance of lactobacilli in centenarians did not differ from that of young adults in the above mentioned Italian cohort.65 However, in a Chinese cohort, the abundance of lactobacilli in rural centenarians (consuming a high fibre diet) was not different from that in healthy urban elderly (consuming a low carbohydrate and low fibre diet), but was lower than that in healthy rural elderly (consuming a high fibre diet).59 Moreover, compared with healthy elderly the abundance of lactobacilli was found to be higher in the subgroup of elderly with CDAD63 70 and in hospitalised elderly.64 However, the CDAD and hospitalised elderly were under metronidazole and undefined antibiotic treatment, respectively, which could have impacted the comparative analyses. van Tongeren et al found that decreased abundance of lactobacilli was associated with deteriorated health status (increased frailty).72 These contrasting observations again highlight the complexity of the impact of chronological age and/or additional host or environmental factors on microbiota composition.
The abundance of streptococci was reported to be mainly higher in healthy elderly and elderly with altered bowel habits compared with young adults,40 64 66 69 although not exclusively.40 None of the studies reported on difference in the abundance of streptococci in centenarians compared with young adults or elderly so far. In addition, the abundance of streptococci was reported to be lower in the subgroup of hospitalised elderly, compared with healthy elderly,64 whereas NSAID use had no effect.69
The family Enterobacteriaceae has diverse ecological (ie, being able to survive in diverse environments) and metabolic characteristics, and includes many potentially pathogenic microorganisms (eg, members of the genera Escherichia, Salmonella, Klebsiella, Proteus). The abundance of Enterobacteriaceae was reported to be higher in faecal samples of healthy elderly than that of young adults from different countries.40 64 Remarkably, a decreased abundance was reported in centenarians compared with young adults in an Italian cohort.65 In a Chinese cohort, however, the abundance of Enterobacteriaceae, as well as the abundance of the genus Escherichia, was higher in rural centenarians than in healthy urban elderly, but was not different from that in healthy rural elderly.59 This indicates that, in addition to health status, the living situation (eg, rural vs urban with differences in, for example, dietary intake and hygiene or antigen exposure) might also contribute to the alterations of intestinal microbiota composition.59 In addition, although the abundance of Enterobacteriaceae was reported to be not significantly different between healthy elderly and hospitalised elderly,73 a higher abundance of Enterobacteriaceae was found in the subgroup of elderly carrying C. difficile compared with C. difficile-negative elderly.74 Moreover, increased Enterobacteriaceae abundance was associated with increased frailty.53 72 This supports the observation that alterations in the intestinal microbiota composition are more pronounced in frail or comorbid elderly.
The observation of our comparative analyses based on the groups identified as ageing-associated by Mitsuoka,62 indicates that in comparison to young adults, the intestinal microbiota of elderly comprised lower levels of bifidobacteria and higher levels of streptococci and Enterobacteriaceae, which seem to be more pronounced in frail or comorbid elderly. However, it should be noted that contradicting findings have also been reported, such as bifidobacteria levels in centenarians. In line with the above observations, a detailed comparative analysis between different studies for other potential microbial taxa associated with elderly or relevant subgroups did not reveal a single microbial group that was consistently positively or negatively associated with ageing (see online supplementary table S1). Moreover, studies that included centenarians or different frailty phenotypes (table 2) hint towards the observation that a decline in health status rather than ageing itself is associated with changes in intestinal microbiota composition. Comparative analyses are further hampered by the myriad of methods38 75 76 used to study the faecal microbiota composition as well as the lack of consistent definitions of frailty53 55 61 72 77 and age for elderly,40 43–45 59 64 72 which could be 80+ years,59 70+ years,44 72 65+ years43 45 64 or 60+ years.40 This stresses the need for well-designed longitudinal studies. Such studies monitoring intestinal microbiota changes over time and also taking into account the large interindividual variation,45 will provide the ideal setting to study intestinal microbiota dynamics during ageing. These studies are obviously long-lasting, adding to complexity, solid scientific infrastructure and costs. Nevertheless, several large-scale population cohorts have already been initiated.67 68 Follow-up of such cohorts may ultimately provide insight into long-term intestinal microbiota dynamics and their relation to ageing, frailty and comorbid conditions.
Overall, we clearly observed that intestinal microbiota compositional changes during ageing are more likely to be associated with health status of the elderly and confounding factors than with ageing itself. This is summarised in figure 1 and table 2 (see also online supplementary table S1 for details per study). It has to be taken into account that most studies focusing on the intestinal microbiota in elderly describe the faecal microbial composition based on 16S rRNA genes. Given the high level of functional redundancy within and across microbial groups as well as the fact that bacteria are very versatile and can quickly adapt and respond to changes in their environment, indicates that only considering composition of the faecal microbiota has its limitations.78 So far, studies focusing on the metabolic capacity or activity of the intestinal microbiota in elderly via metagenomics, metabolomics or other activity-based profiling approaches, are very limited. One study reported that in comparison to young adults, the faecal microbiota of non-institutionalised elderly showed an increase in proteolytic potential, but decreased saccharolytic potential with a low abundance of genes encoding steps in short-chain fatty acids production pathways.41 In line with this observation, decreased concentrations of acetate and propionate, but increased faecal dry matter content and concentrations of branched-chain fatty acids (ie, isovaleric acid, isobutyric acid) have been reported in institutionalised and non-institutionalised elderly compared with young adults.79 80 Furthermore, the living situation of elderly was shown to correlate with the faecal metabolites profile, with higher concentrations of acetate, propionate and valerate in community-dwellers.77 A recent study investigated the functional capacity and activity of the faecal microbiota in a large cohort of healthy elderly (308 men, aged 65–81 years) using metagenomics and metatranscriptomics at four time points over 6 months. However, the study did not report on comparisons between specific subgroups of elderly or dietary habits.67 68 It has been reported that short-term dietary changes can have a drastic impact on microbial metabolite production and host physiology without drastically changing faecal microbiota composition.81 Hence, we argue that approaches addressing functional capacity and activity of the faecal microbiota are crucial to further unravel the role of the microbiota in host physiology of the ageing and/or frail population.
Manipulating the intestinal microbiota of elderly
Although a typical microbiota profile of elderly is hard to define, manipulating the intestinal microbiota of elderly and host outcome has been subject of several studies. Supplementation of functional foods like probiotics, prebiotics or synbiotics are nutritional approaches to beneficially alter the microbiota (figure 1). Several studies have been performed on the effect of probiotics in elderly, of which many focus on the risk of infections and immunosenescence. In a systematic review, including 15 randomised clinical trials in 5916 patients with a mean age of 75 years, Wachholz et al did not find significant effects of probiotics on the occurrence and durations of infections, nor on mortality rate, when compared with placebo.82 Also with regard to inflammatory and immunosenescence markers, Calder et al concluded in a recent review that the evidence for the efficacy of probiotics in elderly is limited and/or inconsistent.83 Several elderly studies did, however, show changes in faecal microbiota composition, being most pronounced for increased abundances of bifidobacteria, after use of, for example, Bifidobacterium lactis HN019, Bifidobacterium longum 46 and Bifidobacterium longum 2C or multispecies probiotics.84 Although probiotic use is often considered to be safe, specific safety studies in elderly are still limited and extra caution is warranted, especially in subjects with impaired host defence mechanisms.
Well-known prebiotics, including galacto-oligosaccharides, inulin and fructo-oligosaccharides,85 have often been evaluated for their effect on bowel habits in constipated elderly, and showed an increase in defecation frequency.86 With regard to immune function, a limited number of prebiotic intervention studies have been performed in elderly. Although beneficial effects on specific parameters have been reported,87 no effect was shown on vaccination efficacy.88 In a synbiotic study, Costabile et al observed a significant and more pronounced effect on NK cell activity, microbiota composition and blood lipids in elderly treated with the combination of Lactobacillus rhamnosus GG and soluble corn fibre, when compared with soluble corn fibre alone.89 Furthermore, different prebiotic and synbiotic food supplement studies have been shown to lead to an increased abundance of faecal bifidobacteria and/or lactobacilli in elderly.84 The clinical relevance hereof, without additional effects on health outcome parameters, is still a matter of debate.
It should be noted that effects of probiotic strains, prebiotic compounds and/or combinations thereof, differ and largely depend on the duration of the intervention period, subject population and mechanism84 to be targeted. We are only at the beginning of understanding the association between specific microbes and our health, especially in elderly and their associated comorbidity or frailty. Although several probiotic, prebiotic and synbiotic studies have been performed in adults in general, caution is needed when extrapolating these findings to elderly and subgroups thereof. Therefore, more insights in the exact microbial composition and underlying mechanistic effects are needed to enable more targeted interventions in relevant subgroups.
Several studies have evaluated dietary intake in general, showing for example, changes in macronutrient intake as well as deficiencies in micronutrients,84 but studies targeting the microbiota composition by changing habitual dietary intake in the elderly are still scarce. Current studies are mainly performed in young adults, showing changes in microbiota composition and especially metabolic activity, relatively quickly after major changes in dietary habits.81 90 91 It should be noted, however, that the observed compositional changes did not exceed the interindividual variation. At present, analyses of the Nu-Age dietary intervention study92 are ongoing, investigating the effect of major diet changes (Mediterranean diet, 1-year randomised, single-blind controlled trial) on the intestinal health in 1250 elderly subjects.
Other strategies to manipulate the intestinal microbiota include faecal microbiota transplantation (FMT). The success rate is variable, and largely depends on the disease or disorder to be treated.93 FMT efficacy is most convincing for treating C. difficile infection,93 which has a rather high incidence in the elderly, also because of the high antibiotic use.94 In a specific review focusing on elderly, Cheng et al confirms the efficacy of FMT for C. difficile in this group, but recommends to use this treatment strategy early in disease course to prevent complications.95 Clear criteria for stool donor selection and screening are still warranted, especially in susceptible (frail) elderly given the risk of transplanting concomitant pathogens and or antibiotic resistance genes.
Summary and conclusions
Several small, age-related declines in the physiology of the GI tract have been reported for ageing in general, whereas frailty and impaired health status seem to play a role in the (further) decline (figure 1). Studies on GI physiology and function in subjects with age-related health decline or being resilient to diseases while ageing, for example, centenarians, as well as studies directly linking host function and outcome in elderly to microbiota composition and activity are, however, still scarce. Regarding the intestinal microbiota composition, alterations are more pronounced in frail or comorbid elderly, although age-related changes in the abundance of Bifidobacterium and Enterobacteriaceae have been reported in general. However, a ‘typical’ intestinal microbiota of elderly is hard to define, given the large interindividual differences in the intestinal microbiota of elderly. Moreover, intestinal microbiota of elderly is more likely to be affected by a broad range of potentially confounding factors, such as lifestyle (eg, including diet and smoking), health status, medical treatment (including medication) and living situation rather than by ageing per se. Although, we acknowledge that unravelling causes and consequences will be challenging since possible confounders such as diet will influence both the microbiota and GI physiology. Based on our current knowledge, future longitudinal studies should shift towards investigating the role of GI physiology and intestinal microbiota as well as their dynamics over time in specific well-characterised subgroups of elderly, such as frailty and elderly with a specific health decline, and how these can be modulated by targeted interventions or improvements in lifestyle and living situation.
Acknowledgments
The authors would like to thank Gerben D A Hermes for his help in discussion about this review, and Roel Bogie for his help with the figure.
References
Footnotes
EGZ and DJ contributed equally.
RA and EW contributed equally.
Contributors All authors were involved in and contributed to this manuscript, and have approved for the final submission.
Funding This work was performed in the public-private partnership ’CarboKinetics' coordinated by the Carbohydrate Competence Center (CCC), as part of the NWO-CCC Partnership Programme. CarboKinetics is financed by participating industrial partners Agrifirm Innovation Center BV, Cooperatie AVEBE UA, DSM Food Specialties BV, FrieslandCampina Nederland BV, Nutrition Sciences NV, VanDrie Holding NV and Sensus BV and allowances [ALWCC.2015.3] of The Netherlands Organisation for Scientific Research (NWO).
Competing interests None declared.
Patient consent Not requried.
Provenance and peer review Commissioned; externally peer reviewed.