Article Text

Abstract
Objective: Persistent stress and life events affect the course of ulcerative colitis and irritable bowel syndrome by largely unknown mechanisms. Corticotropin-releasing hormone (CRH) has been implicated as an important mediator of stress-induced abnormalities in intestinal mucosal function in animal models, but to date no studies in human colon have been reported. The aim was to examine the effects of CRH on mucosal barrier function in the human colon and to elucidate the mechanisms involved in CRH-induced hyper-permeability.
Design: Biopsies from 39 volunteers were assessed for macromolecular permeability (horseradish peroxidise (HRP), 51Cr-EDTA), and electrophysiology after CRH challenge in Ussing chambers. The biopsies were examined by electron and confocal microscopy for HRP and CRH receptor localisation, respectively. Moreover, CRH receptor mRNA and protein expression were examined in the human mast cell line, HMC-1.
Results: Mucosal permeability to HRP was increased by CRH (2.8±0.5 pmol/cm2/h) compared to vehicle exposure (1.5±0.4 pmol/cm2/h), p = 0.032, whereas permeability to 51Cr-EDTA and transmucosal electrical resistance were unchanged. The increased permeability to HRP was abolished by α-helical CRH (9-41) (1.3±0.6 pmol/cm2/h) and the mast cell stabiliser, lodoxamide (1.6±0.6 pmol/cm2/h). Electron microscopy showed transcellular passage of HRP through colonocytes. CRH receptor subtypes R1 and R2 were detected in the HMC-1 cell line and in lamina propria mast cells in human colon.
Conclusions: Our results suggest that CRH mediates transcellular uptake of HRP in human colonic mucosa via CRH receptor subtypes R1 and R2 on subepithelial mast cells. CRH-induced macromolecular uptake in human colon mucosa may have implications for stress-related intestinal disorders.
- CRH receptor subtypes
- barrier function
- electron microscopy
- human mast cell line
- intestinal mucosa
Statistics from Altmetric.com
The association between psychological stress and the development of chronic gastrointestinal disease remains a controversial issue. However, in patients with irritable bowel syndrome (IBS) there is a relationship between stress and symptom severity (for review, see Mayer et al1), and stressful life events appear to increase the risk of developing post-infectious IBS.2 Moreover, persistent stress3 and stressful life events4 have been shown to increase the risk for exacerbations of ulcerative colitis, whereas the epidemiological evidence of an association between stress and Crohn’s disease is less clear.5 The possible mechanisms underlying an effect of stress in intestinal diseases are only beginning to be elucidated.
It is established that the mucosal barrier to environmental antigens is of importance in the disease process in inflammatory bowel disease (IBD). Patients with Crohn’s disease exhibit an increased intestinal permeability,6–8 and display strong immune reactions to dietary antigens and bacteria.9 10 Furthermore, in IBS patients increased permeability persisted for several months after gastroenteritis due to post-infectious immune activation,11 and an increased number of intestinal mucosal mast cells has been demonstrated from gut specimens in such patients.12–14 In animal models, there is a clear link between stress, increased permeability, and gut inflammation (for review, see Söderholm and Perdue15). Rodents exposed to acute stress show increased mucosal permeability to macromolecules via mechanisms that mainly involve corticotropin-releasing hormone (CRH), mast cells and cholinergic nerves.16 17 Chronic stress induces a further increase in permeability, bacterial attachment/penetration, mast cell hyperplasia and inflammatory cell infiltration.18 19 Other studies have revealed that stress can exacerbate mucosal inflammation.20
CRH has been implicated as one of the most important mediators of stress-induced abnormalities in mucosal function in animal models. Stress-induced changes in rat colonic epithelial function can be inhibited by the non-selective CRH antagonist, α-helical CRH (9-41).21–23 In addition, injection of CRH mimics stress-induced changes in colonic function regarding mucin release,16 ion secretion and permeability (paracellular as well as transcellular).21 22 Two main subtypes of CRH receptors, CRH-receptor 1 and 2, have been described. Both receptor subtypes have been found to be expressed in isolated lamina propria mononuclear cells from human colonic mucosa but the exact cell types expressing CRH R1 and R2 have not been determined.24 Selective CRH-R1 antagonists have been shown to inhibit stress effects on intestinal motor function in rats, whereas blocking of the CRH-R2 did not.25 Furthermore, a recent report from Barreau et al showed that in adult rats previously exposed to neonatal maternal deprivation, CRH promoted colonic mast cell release of nerve growth factor (NGF) via the CRH R1 receptor and as a result, gut paracellular permeability was increased.26 However, to date, the relative importance of CRH receptors in human intestinal mucosal function has not been reported.
Our aim in the present study was to examine the peripheral effects of CRH on mucosal barrier function in the human colon. Endoscopic biopsies of normal human colon were exposed to CRH in Ussing chambers and macromolecular permeability was assessed. Further, the mechanisms involved in the CRH-induced hyper-permeability were studied. We found that uptake of macromolecules through colonocytes was indirectly regulated by CRH via activation of CRH-R1 and R2 receptors expressed on subepithelial mast cells.
MATERIALS AND METHODS
Subjects and endoscopy
Thirty-nine healthy volunteers were included, 15 women and 24 men, median age 24 years (range, 20–75). Inclusion criteria were: no medication, no smoking, no history of bowel symptoms and no family history of IBD. Bowel preparation and flexible sigmoidoscopy including biopsies were performed as earlier described.27 The biopsies were immediately put in 4°C oxygenated modified Krebs–Ringer bicarbonate buffer (KRB) and transported to the laboratory within 20 min. Routine clinical histology assessment of sigmoid colon biopsies was normal in all subjects. The study was approved by the Ethical Committee, Faculty of Health Sciences, University Hospital, Linköping, Sweden.
Experimental procedure
All chemicals and drugs were obtained from Sigma Chemical Co., St. Louis, MO, USA unless otherwise indicated.
Studies in human colonic biopsies
Ussing chamber experiments
Biopsies were mounted in modified Ussing chambers (exposed tissue area 1.76 mm2; Harvard Apparatus, Holliston, MA, USA) as previously described and validated by our group.27 Mucosal compartments were filled with 1.5 ml, 10 mmol/l mannitol in KRB and the serosal compartments filled with 10 mmol/l glucose in KRB. The KRB was pH adjusted to 7.4 at 37°C, continuously oxygenated with O2/CO2 (95/5%) and stirred by gas flow in the chambers. After 40 min equilibration to achieve steady state conditions, biopsies with a transepithelial potential difference (PD) less negative than −0.5 mV were excluded due to poor viability.27 Experiments were done in open-circuit conditions with assessment of PD, transmucosal electrical resistance (TER) and short circuit current (Isc) at 2-min intervals using a four-electrode system: Ag/Ag electrodes (Ref 201, Radiometer, Copenhagen, Denmark) with 3 mol/l NaCl/2% agar bridges for PD, and platinum electrodes for current as previously described.28 Samples of 0.3 ml from the serosal side were collected after 0, 30, 60 and 90 min.
Macromolecular permeability
51Cr-EDTA (Perkin Elmer, Boston, MA, USA; mw 342; specific activity of 4662 GBq/mmol (126 Ci/mmol) (3.25 μmol/l) was added on the mucosal side of the tissues and permeation measured by appearance of radioactivity in 0.3-ml serosal samples (counted for 10 min in a 1282 Compugamma reader (LKB, Bromma Sweden)). The apparent permeability coefficient (Papp) was calculated as follows: Papp ( in cm/s) = (dC/dt)V/(Ct0A); where dC/dt is the change in concentration on the submucosal side per unit time (cpm/s), V is the volume of the chamber (cm3), A is the area of exposed colon and Ct0 the initial marker concentration in the mucosal reservoir (cpm).29
Horseradish peroxidase type VI (HRP, mw 45 kDa, 10 μmol/l) was added on the mucosal side. Serosal samples were analysed with QuantaBlu™ Fluorgenic Peroxidase Substrate Kit (Pierce, Rockford, USA) as previously described.30
Dose–response and effects of CRH In the first set of experiments, biopsies were exposed to CRH at 10−8, 10−7 or 10−6 mol/l on the serosal side. Based on dose–response effects, CRH 10−6 mol/l was used in all subsequent experiments.
Effects of CRH receptor blocking agents
In the second set of experiments, after a 20-min equilibration period, the non-specific CRH receptor antagonist, α-helical CRH (9-41), the CRH receptor R1 specific antagonist, astressin, or the CRH receptor R2 specific blocker, antalarmin, was added serosally. For all CRH receptor antagonists, a concentration of 10−6 mol/l was used, which in several studies has been shown to block CRH effects in vitro.31–33 After another 20-min period, CRH was added on the serosal side of tissues mounted in the Ussing chamber.
Effects of mast cell stabilisation
In the third set of experiments, the mast cell stabilising compound, Lodoxamide tromethamine (10−5 mol/l)34 (Alcon Laboratories R&D, Fort Worth, TX, USA) was added 20 min before CRH on the serosal side.
Immunohistochemistry
Biopsies were snap frozen in liquid nitrogen. Cryosections were prepared and fixed with cold acetone. Sections were incubated with rabbit anti-human tryptase polyclonal antibody (1 μg/ml, Serotec, Toronto, ON, Canada) and goat anti-human CRH R1 or R2 (1 μg/ml for both CRH R1 and R2; Santa Cruz, CA, USA) for 1 h at room temperature. After washing, sections were incubated with Cy5-anti-goat second antibody (50 ng/ml, DAKO, Oakville, ON, Canada) and FITC-anti-rabbit second antibody (50 ng/ml, DAKO, Oakville, ON, Canada) mixed with propidium iodide (10 μg/ml, Sigma, Oakville, ON, Canada) for 1 h. In a second set, serial sections of the biopsies were performed. Sections were stained for CRH R1 and R2 as described above except that FITC anti-goat secondary antibody was used (50 ng/ml, DAKO, Oakville, ON, Canada). Sections were observed with confocal microscopy.
Transmission electron microscopy
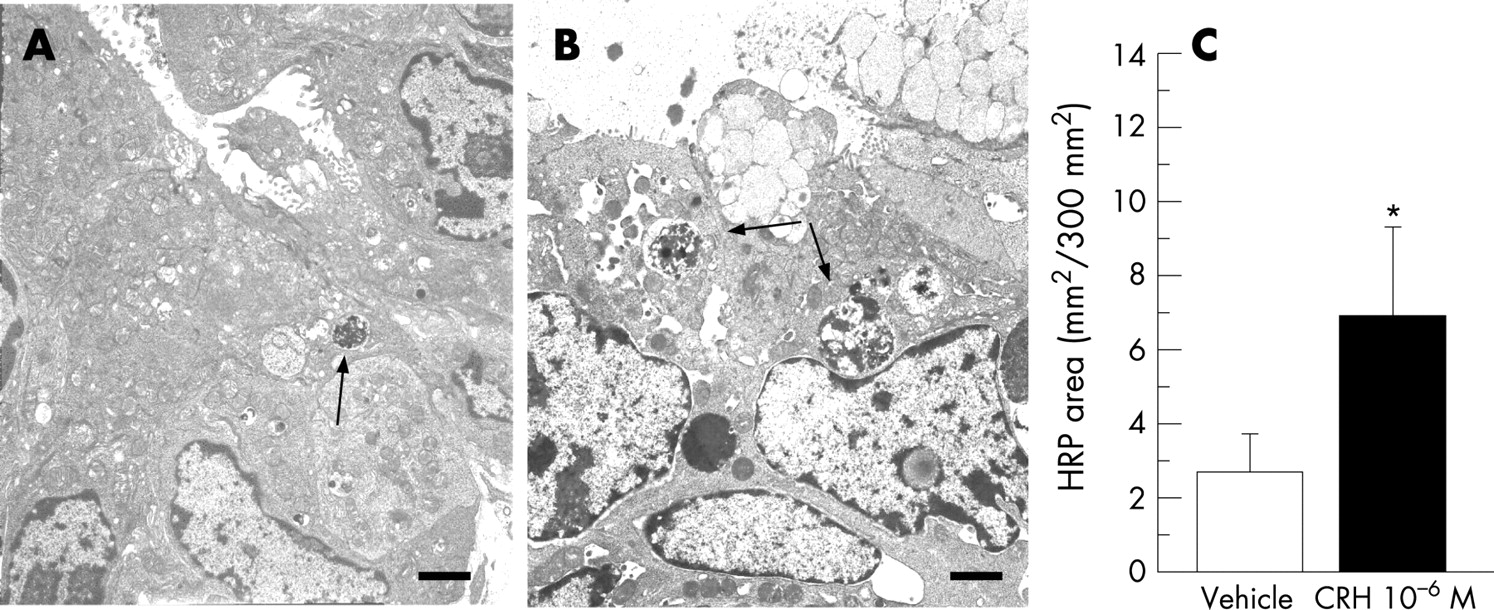
Tissues were fixed in 2% glutaraldehyde in 0.1 mol/l sodium cacodylate buffer (pH 7.4) for 2 hours at room temperature, washed in several steps, incubated with 5 mg of 3,3′-diaminbenzadine tetrahydrocloride in 10 ml of 0.05 mol/l Tris buffer and 0.01% H2O2 (pH 7.6 at 22°C). HRP product identification was modified from Graham and Karnovsky35 as previously described.36 Quantitative analysis of HRP uptake in intracellular endosomes was performed on coded high magnification photomicrographs from the surface epithelium. HRP containing endosomes within a fixed size observation window (300 μm2) in the apical region of the colonocytes were counted and measured using a computerised image analysis system (Kontron Mop Videoplan, Kontron, Eching, Germany). Twenty photomicrographs from each treatment were examined in biopsies from three volunteers, with investigator unaware of CRH treatment group. The area of HRP-containing endosomes was calculated and averaged for each volunteer before determining group means. Mast cells were evaluated for signs of activation by image analysis. Patterns of mast cell activation/degranulation were assessed as piecemeal degranulation, defined as the loss of granule density without granule-cell membrane fusion,19 or anaphylactic degranulation, defined as solubilisation of granular contents with fusion of inter-granular and granule-cell membranes.20 Sections from each tissue were randomly selected, 30 mast cells were identified on each grid, analysed for loss of density and perigranular vacuolisation, and classified as activated or non-activated.
Studies in the human mast cell line HMC-1
Cell culture
The human mast cell line, HMC-1 (a gift from Dr J.H. Butterfield, Mayo Clinic Rochester, USA) was cultured in RPMI 1640 containing 10% iron supplemented FBS, 1.2 mmol/l α-thioglycerol, 100 U/ml penicillin, 10 μg/ml streptomycin (5% CO2, 37°C), and 2 mmol/l glutamine.
Total RNA preparation and cDNA synthesis
HMC-1 cells were centrifuged for 5 min at 500 rpm, cell pellets were washed twice with PBS. Total RNA was prepared from the cell pellets using the RNeasy Total RNA Kit (QIAGEN, Mississauga, ON, Canada). First strand cDNA synthesis was performed using ribonuclease reverse transcriptase (Life Technologies, Inc., Mississauga, ON, Canada).
RT-PCR
All PCR reactions were carried out using Taq DNA polymerase (Perkin-Elmer, Mississauga, ON, Canada) with 200 ng cDNA for each amplification, as previously described.36 Primers of the subtypes of CRH receptor 1 and 2 using for amplification were designed with the software Primer3. The specificity of the primers was confirmed by executing the software Blast. CRH-R1 primer (NM_004382, 267 bp): sense, 5′- TCTCAGGACTGCAGTGCAAC-3′ and antisense, 5′-AGTGGCCCAGGTAGTTGATG-3′; CRH-R2 primer (NM_001883, 240 bp). Sense, 5′-TTCCAGGGTTTCTTCGTGTC-3′; anti-sense, 5′- GGAAGAAGGTGGAGGAGGAC-3′. human glyceraldehyde-3-phosphate dehydrogenase (GADPH) primer (NM_002046, 238 bp). Sense, 5′-GAGTCAACGGATTTGGTCGT-3′; anti-sense, 5′-TTGATTTTGGAGGGATCTCG-3′. The cycle program for: CRH-R1 primers consisted of 35 runs of denaturation at 94°C for 30 s, annealing at 62°C for 30 s, and elongation at 72°C for 30 s; CRH-R2 primers consisted of 40 runs of denaturation at 94°C for 1 min, annealing at 55°C for 1 min, and elongation at 72°C for 1 min; GADPH primers consisted of 25 runs of denaturation at 94°C for 1 min, annealing at 56°C for 1 min, and elongation at 72°C for 1 min. The cycle programme was preceded by an initial denaturation at 94°C for 3 min and followed by a final extension at 72°C for 10 min. Electrophoresis was subsequently done with 20 μl of the reaction mixture on a 1.2% agarose gel and visualised by ethidium bromide, using a 1-kb DNA ladder (Perkin-Elmer, Mississauga, ON, Canada) to estimate the band sizes. As a negative control for all of the reactions, distilled water was used in place of the cDNA.
In situ detection of CRH receptor mRNA
Oligo-probes were synthesised by Mobix Biolab (McMaster University, Hamilton, ON, Canada) with FITC tags, with sense strands of the oligo sequences as controls. HMC-1 cells were harvested, fixed with absolute methanol for 10 min, rehydrated, and stored in PBS at 4°C. Probes for CRH R1, R2 and control were: Oligo-1 (CRF-R2, antisense): (5′-fluo-TCCGGGTCGTGTTGTACTTGACGCCGTTGAAGTACTCGGGGCACG-3′). Oligo-2 (CRF-R2, sense): (5′-fluo-CGTGCCCCGAGTACTTCAACGGCGTCAAGTACAACACGACCCGGA-3′) and Oligo-3 (CRF-R1, antisense); (5′-fluo-CCACTAGCTGCCCCGCAGGGCT GCGGGGCCAGCAGGTGCC-3′) For in situ hybridisation fixed HMC-1 cells were smeared on Aptex coated slides and dried at room temperature overnight. All slides were treated with pre-hybridisation solution for 30 min, followed by adding hybridisation solution with oligo-probes or their controls overnight at 37°C. Slides were then placed in preheated (45°C) 2× standard saline citrate buffer, in which they were washed twice, followed by another 10-min immersion in 0.1× standard saline citrate (45°C), rinsed with PBS for 5 min, mounted with coverslips and observed by confocal microscopy.
Western blot
HMC-1 cells were lysed and proteins quantified spectrophotometrically using Bio-Rad DC Protein Assay kit (CA, USA). Equivalent amounts (40 μg) of total cellular protein were fractionated by SDS-PAGE and blotted onto nitrocellulose membranes (Amersham Biosciences, Oakville, Ontario, Canada). Membranes were sequentially incubated with primary antibody to CRH receptor 1 or CRH receptor 2 (goat anti-human CRH receptor 1 or 2, diluted 1/10000). HRP-conjugated rabbit anti-goat secondary antibody (1/5000; the two receptors were detected separately). Chemiluminescence was detected by using SuperSignal West Pico Chemluminescent Substrate (Pierce, Rockford, IL). Ovalbumin was used as negative size matched control.
Immunocytochemistry
HMC-1 cells were fixed in 4% formaldehyde for 30 min, washed with 0.05 mol/l Tris buffer, and incubated for 2 h in PBS with 5% normal goat serum. Goat anti-human CRH receptor 1 or 2 antibodies (Santa Cruz, USA), respectively, were diluted 1:100 in 1% BSA-PBS containing 0.1% Triton-X 100. Cells were incubated for 1 h with FITC-conjugated rabbit anti-goat antibody (Vector Laboratories, Burlingame, CA) diluted 1:200 in 1% BSA-PBS, and 5 μg/ml propidium iodide. After washing, cells were smeared on glass slides, covered in antifade medium (Vectashield; Vector Laboratories), and observed by confocal microscopy. Negative controls omitted the primary antibodies or were incubated with a non-specific isotype-matched IgG.
Flow cytometry
The HMC-1 cells were processed as described above. After staining, the positively stained cells were counted with a FACScan. At least 10 000 cells were analysed for each sample using the CellQuest and WinMDI.
Statistics
Data are presented as means ± SEM. In Ussing chamber experiments, the n value represents the number of volunteers, with a mean value for each volunteer calculated from two to three biopsies for each treatment. Comparisons between treatment groups were done with two-way ANOVA with post-hoc tests and the Student t test as applicable, and comparisons of paired data over time were assessed with the paired t test; p<0.05 was considered as significant.
RESULTS
CRH increased macromolecular transport in human colon
Mucosal permeability to HRP during the 30–90 min period is outlined in fig. 1. There was an increased permeation of HRP when the serosal side of biopsies were exposed to CRH. The HRP transport showed a concentration–response pattern, with significantly increased permeability compared to controls at a CRH concentration of 10−6 mol/l, (p = 0.0068). Therefore, this concentration was used in the subsequent experiments. By electron microscopy, an augmented endocytotic uptake of HRP was shown by the increased area of HRP-containing intracellular vesicles compared to controls, fig. 2.
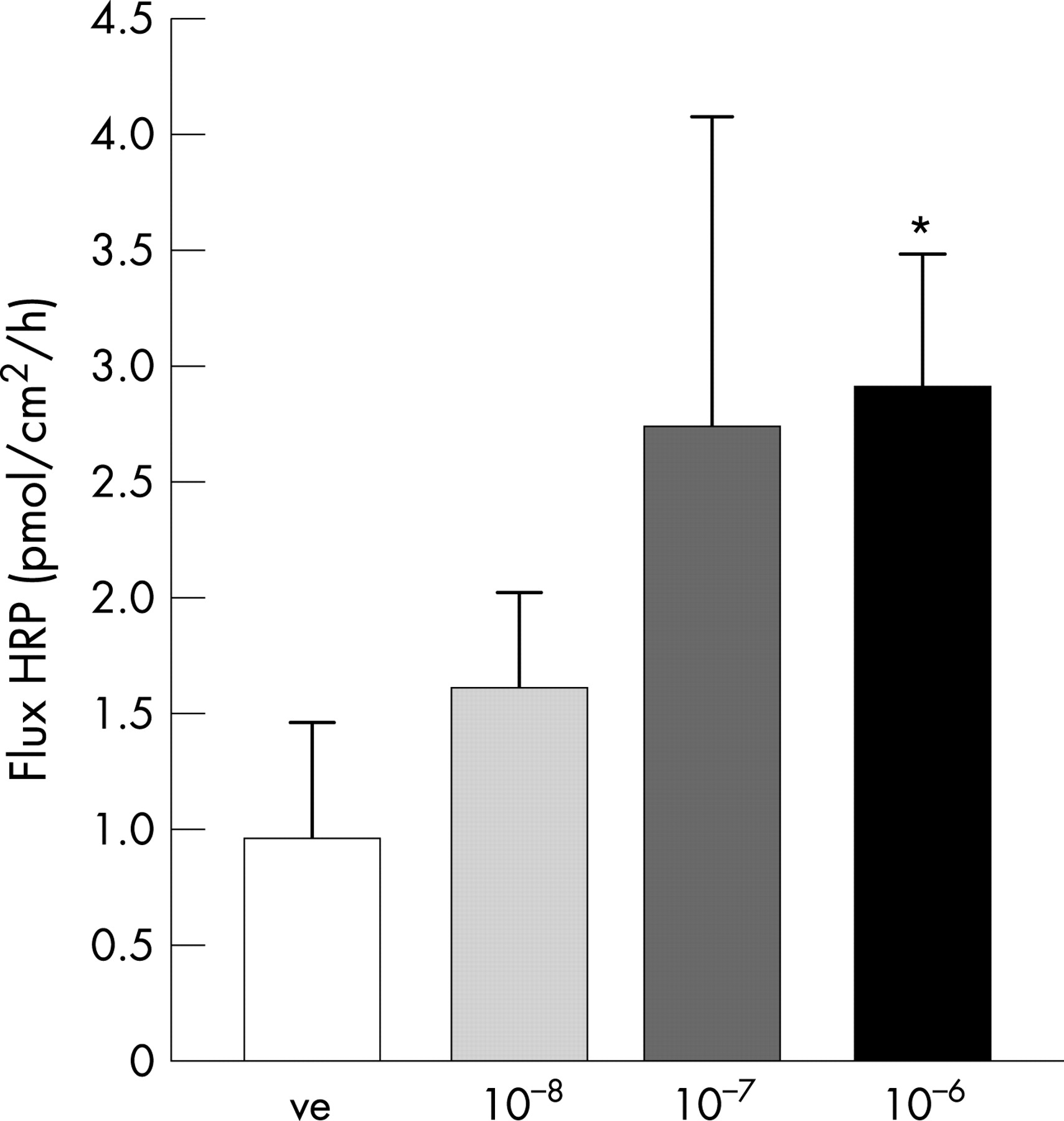
There was no increase in the permeation of 51Cr-EDTA during exposure to CRH (Papp; 1.46±0.09×10−6 cm/s) compared to controls (Papp; 1.47±0.07×10−6 cm/s) during the 30–90 min period; n = 34 volunteers. In addition, there was no effect of CRH on TER or Isc (fig. 3).

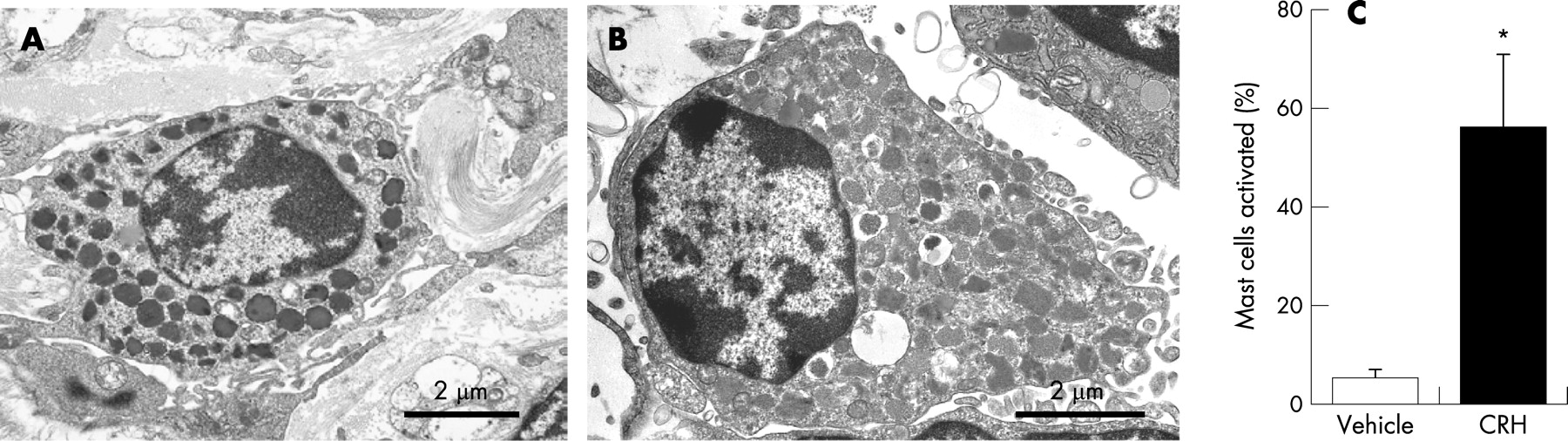
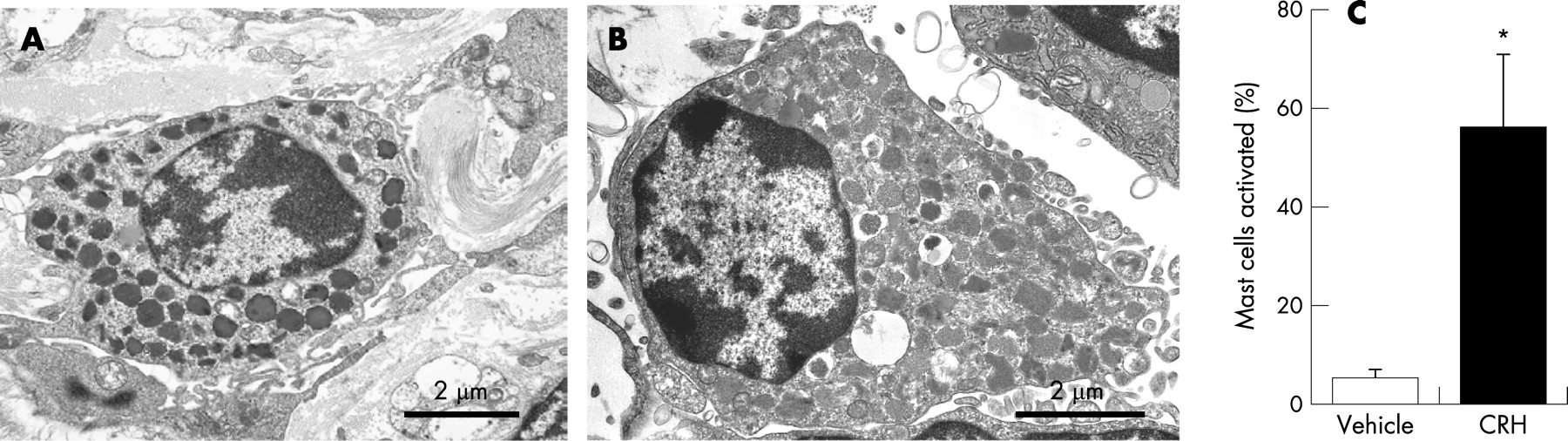
CRH activated mast cells
The numbers of activated versus non-activated mast cells was analysed by using electron microscopy. There was a significant difference in activation of mast cells between the biopsies exposed to CRH and those exposed to vehicle (fig. 4). Further proof of mast cell activation by CRH was seen when the specimens were pre-treated with lodoxamide [10 μmol/l] 20 min before CRH was added. The increased HRP uptake during CRH exposure was abolished by pre-treatment with lodoxamide (2.8±0.5 vs. 1.6±0.6 pmol/cm2/h) (fig. 5).

CRH R1 and R2 were expressed in human mast cell line, HMC-1
To determine if human mast cells express CRH receptors, mRNA and protein expression of the two receptor subtypes CRH-R1 and R2 were assessed in the human mast cell line, HMC-1. RT-PCR (fig. 6a) and in situ hybridisation (fig. 6b–d) demonstrated that mRNA for both receptor subtypes was expressed in HMC-1 cells. These findings were confirmed at the protein level by western blot (fig. 7). In addition, immunocytochemistry demonstrated cell membrane expression of the receptors in HMC-1 cells (fig. 7). Moreover, protein expression was quantified by FACS scan analysis, detecting both CRH-R1 and CRH-R2 in approximately 40% of HMC-1 cells.


CRH R1 and R2 were expressed in human colonic mast cells
In human colon specimens CRH R1 as well as R2 receptors were detected by immunohistochemistry. In all sections, expression was localised in lamina propria cells close to the epithelial layer. With double staining for mast cell tryptase and CRH-R1 or CRH-R2, these receptors were found to be expressed exclusively, but not on all, mast cells. Moreover, in serial sections, mast cells were noted to express both CRH receptors (fig. 8).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
No difference between CRH-R1 and R2 in regulation of HRP uptake
In order to evaluate the functional importance of the receptor subtypes in the regulation of HRP permeation, the CRH-R1 antagonist, astressin, and the CRH-R2 antagonist, antalarmin, and the non-specific antagonist, α-helical CRH (9-41), were used. The increased flux of HRP in CRH exposed tissue (2.8±0.5 pmol/cm2/h) was abolished by pre-treatment with the non-selective CRH receptor antagonist α-helical CRH (9-41) (1.3±0.6 pmol/cm2/h) (fig. 5). The CRH receptor 1 antagonist, astressin (2.0±0.15 pmol/cm2/h), and the CRH receptor 2 antagonist, antalarmin (1.8±0.4 pmol/cm2/h), also resulted in partial inhibition, and were not different from control (1.5±0.4 pmol/cm2/h). Fluxes of HRP were measured during the 30–90 min period.
DISCUSSION
In this first study of the functional effects of the putative stress mediator, CRH, in human intestinal mucosa, we found that CRH activated subepithelial mast cells and increased transcellular uptake of protein antigens in human colonic mucosa. CRH receptors R1 and R2 were expressed on mast cells and CRH-receptor antagonists inhibited the permeability increase evoked by CRH. Our findings suggest that CRH, acting via mast cells, is involved in regulating macromolecular permeability in the normal human colon.
Transmission electron microscopy demonstrated that CRH stimulated mast cells, which was corroborated by the inhibiting effects of pre-treatment with lodoxamide. This mast cell stabiliser exerts its effect by blocking Ca2+ flux into the mast cell, thereby inhibiting activation and degranulation.37 Electron microscopy also showed that the increased HRP permeability was an effect of increased transcellular transport via endosomes. These findings are in agreement with studies on CRH effects in rat intestine.21 Partly contradictory to previous studies, we found paracellular pathways to be unaffected by CRH, with unchanged permeability to the paracellular probe, 51Cr-EDTA, and to ions as assessed by TER. In addition, Isc was unaffected by CRH. Crowe et al reported that when mast cells in human colonic specimens were activated by anti-IgE serum, a histamine-dependent increase in Isc was found.38 Furthermore, Jacob et al reported decreased TER, redistribution of tight junction proteins, and increased transepithelial flux of macromolecules mediated by mast cell tryptase release in T84 cells after exposure to mast cells activated by compound 48/80.39 Ferrier et al showed increased colonic paracellular permeability to 51Cr-EDTA in a murine model of acoustic and restraint stress. However, the increased permeability could not be explained by a direct effect of CRH alone, but was dependent of IFN-gamma production and CD4+ cells.40 Other recent reports suggest that mast cells can show a more selective degranulation pattern. For example, in the HMC-1 cell line, exposure to CRH induced a selective release of vascular endothelial growth factor (VEGF) which could be blocked by the CRH-R1 antagonist, antalarmin.33 Moreover, Barreau et al showed, in the neonatal maternal deprivation model, that NGF is released from colonic mast cells in response to CRH via the CRH-R1 receptor.26 In the present study, CRH exposure did not induce anaphylactic-type degranulation of colonic mucosal mast cells (with release of histamine and tryptase), but generally caused a piece-meal type degranulation pattern, in accordance with previous observations.36 Therefore, mast cells may release compounds specifically acting on epithelial cells to increase endosomal transport of antigens without causing changes in tight junction permeability and ion transport. In theory, TNFα, a preformed mast cell factor which in low concentrations has been shown to increase transcellular protein uptake without affecting paracellular permeability in human mucosa and epithelial cells,41 could be released by CRH exposure. Further studies of selective mast cell activation in the gastro-intestinal tract are needed to elucidate factors that mediate the increased transcytosis.
CRH receptor expression was clearly demonstrated on the mast cells in the HMC-1 line. We found mRNA for CRH-R1 and R2 together with expression of the receptor subtype proteins located at the cell membranes. In human colonic mucosa, we also found CRH R1 and R2 receptor expression on mast cells. Moreover, serial sections suggested that many mast cells expressing CRH receptors expressed both subtypes. Using double-staining with mast cell tryptase, the CRH receptors were identified exclusively on mast cells in the lamina propria, although not on all mast cells. Since there are several animal studies describing CRH receptors on colonic goblet cells, stem cells, myenteric plexus neurons, and macrophages, in addition to mast cells, our present findings were somewhat surprising. As an explanation, firstly, the colonic biopsies studied did not include the myenteric plexus. Further, inflammation has been described to induce CRH receptor expression,42 whereas the volunteers in this study were all healthy, with no pathology being observed on routine clinical light microscopy assessment. Most importantly, in the only previous report on CRH receptor subtypes in human colonic mucosa, Muramatsu et al found CRH receptors to be localised to non-specified lamina propria mononuclear cells,24 which may well have been mast cells. Thus, our finding of CRH receptor expression only on mast cells in the lamina propria of the human colon is in line with previous evidence in humans.
We were also able to demonstrate functionality of the CRH receptors in the colonic biopsies, with a clear dose–response pattern to CRH exposure, and abolished permeability effects when adding the non-selective CRH receptor blocker, α-helical CRH (9-41). In the central nervous system, the CRH receptor subtypes have been found to exert distinct functions, sometimes opposing each other.43 44 Whereas CRH R1 is involved in stress related responses and visceral pain, activation of CRH R2 dampens the behavioural stress responses.45 46 Recently it was also shown that CRH R2 has a role in regulating the inflammatory response in the ileum of mice,47 with a reduced response to Clostridium difficile toxin in CRH R2-null mice. There is so far no description of the function of CRH-R1 and CRH-R2 receptors on human colonic mast cells. Furthermore, different isoforms of the CRH- R1 and R2 receptor may be of importance. In a report from Cao et al expressions of both mRNA and protein for the isoforms CRH R1α, R1β, R1c, R1e, R1f, CRH R2α, R2β and R2γ were investigated in the HMC-1 cell-line and in human umbilical cord blood-derived mast cells (h-CBMC). The functional significance of the individual CRH receptor isoforms in mast cells is still unknown.33 Our findings suggest that both receptor subtypes are involved in the regulation of the mucosal barrier, emphasising the need to further elucidate the CRH–CRH receptor–mast cell–epithelial cell cross-talk.
In vitro techniques, like Ussing chambers, have limitations (for example, the lack of circulation and nervous control) but it is necessary to use this approach for studies of mechanisms involved in human intestinal mucosal function. In a previous study to evaluate our technique,27 we found that human intestinal biopsies have good viability in Ussing chambers, and can be used to study transcellular uptake of protein antigens and paracellular permeability to marker molecules. Moreover, the study showed that mucosal permeability as well as electrophysiological parameters can detect barrier alterations in our system.27 Another issue is the broad distribution of data in the control group, which is higher than usually seen in experimental studies in animals. The present study was, however, done in a consecutive group of volunteering human subjects with very diverse genetic and environmental backgrounds. In spite of this, we did find significant CRH effects on HRP uptake. Although one must use caution in extrapolating our findings to the complex in vivo situation in the human intestine, we believe that our data provide evidence that CRH and mast cells are involved in the regulation of barrier function in the human colon. Given the importance of CRH and mast cells in animal models of stress and intestinal inflammation,15 and their potential role in IBS13 48 and IBD,49 50 a pertinent next step is to study changes in this signalling system in various patient groups.
In conclusion, we present evidence that CRH, an important neuroendocrine mediator released during stress, activates CRH receptor subtypes R1 and R2 on subepithelial mast cells, which in turn stimulate increased transcellular uptake of antigen-sized macromolecules in human colonic mucosa. Since increased uptake of antigenic molecules is associated with mucosal inflammation, our findings may have implications for understanding stress-related intestinal disorders such as IBS and IBD.
Acknowledgments
The authors wish to thank Mrs Ylva Braaf for skilful technical assistance.
REFERENCES
Footnotes
Funding: This study was supported by the Swedish Research Council (VR-M), the Swedish Society of Medicine (Ihre Foundation), the Crohn’s and Colitis Foundation of Canada (CCFC) and the Canadian Institutes for Health Research (CIHR). DMM holds a Canada Research Chair in Intestinal Immunophysiology and is supported by the Alberta Heritage Foundation for Medical Research (AHFMR). PMS is the recipient of a Canada Research Chair in Gastrointestinal Disease.
Competing interests: None.