Article Text

Abstract
Background: Transforming growth factors βs (TGF-βs) are implicated in pancreatic tissue repair but their role in acute pancreatitis is not known. To determine whether endogenous TGF-βs modulate the course of caerulein induced acute pancreatitis, caerulein was administered to wild-type (FVB−/−) and transgenic mice that are heterozygous (FVB+/−) for expression of a dominant negative type II TGF-β receptor.
Methods: After 7 hourly supramaximal injections of caerulein, the pancreas was evaluated histologically and serum was assayed for amylase and lipase levels. Next, the effects of caerulein on amylase secretion were determined in mouse pancreatic acini, and cholecystokinin (CCK) receptor expression was assessed.
Results: The normal mouse pancreas was devoid of inflammatory cells whereas the pancreas from transgenic mice contained lymphocytic infiltrates. Caerulein injection in wild-type mice resulted in 6- and 36-fold increases in serum amylase and lipase levels, respectively, increased serum trypsinogen activation peptide (TAP) levels, gross oedema and a marked inflammatory response in the pancreas that consisted mainly of neutrophils and macrophages. By contrast, FVB+/− mice exhibited minimal alterations in response to caerulein with attenuated neutrophil–macrophage infiltrates. Moreover, acini from FVB+/− mice did not exhibit restricted stimulation at high caerulein concentrations, even though CCK receptor mRNA levels were not decreased.
Conclusion: Our findings indicate that a functional TGF-β signalling pathway may be required for caerulein to induce acute pancreatitis and for the CCK receptor to induce acinar cell damage at high ligand concentrations. Our results also support the concept that restricted stimulation at high caerulein concentrations contributes to the ability of caerulein to induce acute pancreatitis.
- CCK, cholecystokinin
- LDH, lactate dehydrogenase
- TAP, trypsinogen activation peptide
- TGF, transforming growth factor
- TβR, TGF-β receptor
Statistics from Altmetric.com
- CCK, cholecystokinin
- LDH, lactate dehydrogenase
- TAP, trypsinogen activation peptide
- TGF, transforming growth factor
- TβR, TGF-β receptor
Acute pancreatitis has a wide range of clinical symptoms that vary from mild, self-limiting upper abdominal pain to multiorgan failure and sepsis with high mortality. Several aetiologies are known to induce acute pancreatitis, including biliary tract stones, ethanol abuse, trauma, tumours and idiopathic factors. The most frequent causes for acute pancreatitis are gallstone disease and alcohol consumption in Western countries.1
Caerulein induced acute pancreatitis is one of the best studied animal models for this disease.2,3 Caerulein normally causes a mild, necrotising pancreatitis in mice which is self-limiting and highly reproducible. It is thought that the initiation of this disease is mediated through premature intracellular activation of zymogens in the acinar cells. This leads to activation of a cascade of digestive zymogens which finally results in acinar cell necrosis and mild pancreatic oedema, and is followed by an inflammatory response.4,5 Furthermore, amylase leakage from injured acinar cells causes hyperamylasaemia.4,5
Transforming growth factors βs (TGF-βs) and their receptors (TβR) belong to a large family of cytokines which are involved in the immune and inflammatory response, tissue repair, and cell growth and differentiation.6 They play an important role in pancreatic repair after acute pancreatitis and may contribute to the development of chronic pancreatitis.7,8,9,10 Thus TGF-β expression is increased after induction of acute pancreatitis in rats,9 TGF-βs promote the development of pancreatic fibrosis after repeated courses of caerulein induced acute pancreatitis8 and TGF-βs regulate the remodelling of extracellular matrix during regeneration.7 TGF-βs may also regulate neutrophil function, and pancreatic neutrophil infiltration may be responsible in part for acinar cell necrosis.11
Transgenic mice expressing a dominant negative TβRII in a heterozygous manner (FBV+/-) exhibit functional inactivation of the growth inhibitory actions of all three TGF-β isoforms in pancreatic acinar cells in conjunction with increased acinar cell proliferation, altered acinar differentiation, acinar cell apoptosis, neoangiogenesis, mild macrophage and mast cell infiltration, and mild pancreatic fibrosis12 (our own data). These morphological changes increase progressively with age but are not associated with overt signs of exocrine pancreatic insufficiency such as weight loss. To assess the potential role of TGF-βs in the early stages of acute pancreatitis, in the present study we took advantage of the availability of this transgene model. We now demonstrate that mice lacking a functional TβRII do not develop acute pancreatitis following caerulein administration. We also demonstrate that pancreatic acini from these mice fail to exhibit cellular damage or restricted stimulation of secretion in response to high concentrations of caerulein. These results point to interactions between the TGF-β and cholecystokinin (CCK) receptor dependent pathways, and raise the possibility that signalling through TGF-β receptors may contribute, directly or indirectly, to acinar cell damage induced by excessive stimulation of the CCK receptor.
MATERIALS AND METHODS
Animals and reagents
FVB/N mice were engineered to express a human dominant negative TβRII consisting of the extracellular and transmembrane domains of the receptor.12 Dominant negative TβRII expression was under the control of a modified metallothionein promoter, thereby generating hemizygous AM3 mice (termed FBV+/−), as previously described.12 These FBV+/− mice express 3 copies of the truncated receptor per haploid genome.12 The transgenic mice did not suffer any immune alterations, were not prone to infections and did not require special housing precautions. In this regard it should be noted that the metallothionein 1 promoter allowed for expression of the dominant negative TβRII in the pancreas and liver due to the abundance of zinc in these organs. By contrast, neither the spleen nor the thymus exhibited expression of the transcript, even when the mice were administered a large dose of zinc.12
Restriction enzymes and the random primed labelling kit were from Boehringer-Mannheim (Indianapolis, Indiana, USA); [α-32P]dCTP was from Amersham (Arlington Heights, Illinois, USA); Genescreen membranes were from New England Nuclear (Boston, Massachusetts, USA); trypsinogen activation peptide (TAP) enzyme immunoassay kit from Biotrin International (Sinsheim-Reihen, Germany); lactate dehydrogenase (LDH) assay kit from PanVera Corporation (Madison, Wisconsin, USA); and purified collagenase (CLSPA) from Worthington Biochemicals (Lakewood, New Jersey, USA). Caerulein and all other chemicals and reagents were from Sigma Chemical Co. (St. Louis, Missouri, USA).
Experimental procedures
Pancreatitis was induced by seven subcutaneous injections of caerulein (50 μg/kg body weight) given at hourly intervals, based on subcutaneous injection models that have been previously described.13–15 Mice from the control groups received subcutaneous injection of carrier buffer (6% dextran 70, diluted in 0.9% NaCl). All animals were fasted for 12 hours before the experiment and given access to water ad libitum. In total, four groups of animals were evaluated: FBV−/− sham or caerulein-injected, and FBV+/− sham or caerulein-injected. Every group consisted of six animals per group. The animals were sacrificed 16 h after the last injection of caerulein. The pancreata were removed and one part was frozen in liquid nitrogen and stored at −80°C for RNA extraction; the other part was fixed in paraformaldehyde and paraffin embedded for histological assessment. All studies with mice were approved by the University of California, Irvine and by Dartmouth Medical School Institutional Animal Care and Use Committees.
Pancreas histology
After staining with H&E, pancreatic slides (4 μm) from caerulein or sham injected mice were graded in a blinded manner, without knowledge of the experimental design, according to the severity and extent of oedema, inflammatory cell infiltration and acinar necrosis,16 as described in table 1.
Criteria used for histological analysis of severity of acute pancreatitis, as described by Van Laetham and colleagues8
RNA extraction and northern blot analysis
Total RNA from the pancreas was extracted by the single step acid guanidinium thiocyanate phenol chloroform method.17 RNA samples from three mice per group were pooled, size fractionated on 1.2% agarose/1.8 M formaldehyde gels, electrotransferred onto nylon membranes and cross linked by UV irradiation. Blots were prehybridised and hybridised with the indicated cDNA probes and washed under high stringency conditions, as previously reported.17 Blots were then exposed at −80°C to Kodak BiomaxMS films. A BamHI 190 kb fragment of mouse 7S cDNA was used to confirm equal RNA loading and transfer.17
Serum amylase, lipase and TAP levels
Blood was collected by cardiac puncture immediately after sacrificing the mice. Following centrifugation, serum was stored at −20°C until assayed. Serum α amylase was measured using 4.6-ethylidene-(G7)-1-4-nitrophenyl-(GI)-α, D-maltoheptaoside as the substrate (EPS method) according the manufacturer’s protocol (Boehringer Mannheim). Lipase was measured using the peroxidase test.18 TAP was assayed by ELISA using an enzyme immunoassay kit according to the manufacturer’s instructions.19
Determination of pancreatic water content
The water content of the pancreas was quantified by comparing the weight of the freshly harvested tissue (wet weight) with the weight of the same sample after desiccation (dry weight). To assure complete desiccation, the tissue was incubated at 90°C for 36 h, by which time it had achieved a constant weight. Results are expressed as the percentage of the difference between the wet weight and dry weight, divided by the original wet weight.20
Amylase secretion studies
Pancreatic acini were prepared as previously reported, using collagenase digestion, followed by mechanical dispersion, and passed through a mesh nylon cloth.21,22 Acini were then purified by centrifugation of 50 g for 1 min in colourless Dulbecco’s modified Eagle’s medium containing 0.2% bovine serum albumin and were resuspended after three washing steps in incubation medium. Acini were allowed to recover for 30 min (23°C) prior to use in experiments. Amylase content was measured in cell pellets after lysis (4°C) with buffer that consisted of 50 mM HEPES, 0.1% Triton X-100, 5 mM EDTA, 150 mM NaCl, 10% glycerine and a protease inhibitor cocktail (1 ml/mg tissue) that consisted of 10 μg/ml aprotinin, 10 μg/ml leupeptin, 0.01 M sodium pyrophosphate, 0.1 M sodium fluoride, 1 mM L-phenyl-methyl-sulfonyl-fluoride and 0.02% SBTI. Amylase release was measured by enzymatic reaction using ET-G7PNP as substrate. The percentage of amylase release from the cells after stimulation was expressed as the ratio of amylase concentration in the supernatant divided by amylase concentration in cell lysates. Viability of pancreatic acini was routinely checked by trypan blue exclusion and only preparations with >95% cellular viability were subjected to secretion studies. In addition, LDH leakage from pancreatic acini cells after stimulation with secretagogues was determined in the supernatant, using a microtitre plate (ELISA) reader (Molecular Devices, Sunnyvale, California, USA), with absorbance set at 490 nm and reference wavelength at 650 nm, and an assay kit from PanVera according to the manufacturer’s instructions.
RESULTS
Expression of TβRII and ligands by northern blot analysis
To confirm the previously reported findings,12 we initially determined levels of expression of the truncated TβRII mRNA in the pancreata of wild-type (FVB−/−) and transgenic (FVB+/−) mice. In FVB+/− mice, a strong TβRII mRNA signal was evident (fig 1), in agreement with the anticipated size of the truncated transcript (5.2 kb). In contrast, the truncated TβRII mRNA signal was not present in the pancreas from wild-type mice (fig 1).
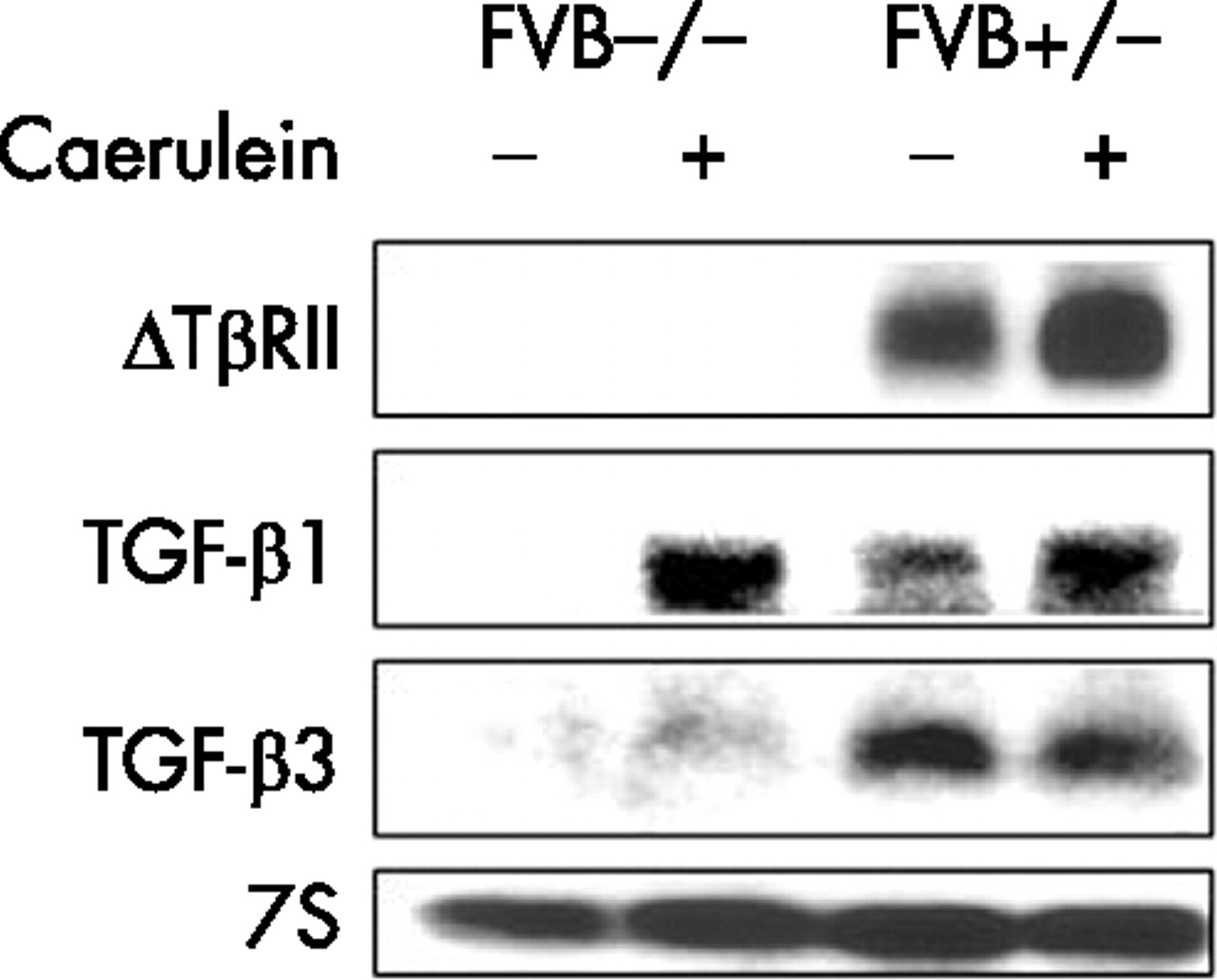
Expression of type II transforming growth factor β receptor (TβRII) and ligands. Total pancreatic RNA (20 μg/lane) was isolated from wild-type (FVB−/−) and transgenic (FVB+/−) mice that were either injected with buffer (sham) or with caerulein, as described in the Methods section. RNA was subjected to northern blot analysis using 32P-labeled cDNA probes (500 000 cpm/ml) specific for truncated TβRII, transforming growth factor (TGF)-β1 and TGF-β3. A 7S ribosomal cDNA probe (50 000 cpm/ml) was used as a loading and transfer control. Exposure times were overnight for TβRII, TGF-β1 and TGF-β3, and 3 h for 7S.
Next, TGF-β1, TGF-β2 and TGF-β3 mRNA levels were determined (fig 1). The TGF-β1 (2.4 kb) mRNA transcript was below the level of detection by northern blot analysis in the pancreas of FVB−/− mice whereas the TGF-β3 mRNA transcript (3.8 kb) was very faint and only clearly visible on the original autoradiograph. Both TGF-β1 and TGF-β3 transcripts were expressed at relatively high levels in the pancreas from FVB+/− mice, as previously reported in this transgene model.12 After caerulein treatment, FVB−/− mice exhibited a marked increase in pancreatic TGF-β1 mRNA levels and a very slight increase in TGF-β3 mRNA levels (fig 1). FVB+/− mice also exhibited an increase in TGF-β1 mRNA levels whereas the high TGF-β3 mRNA levels in the sham injected mice were slightly decreased. Caerulein also caused an increase in the levels of the truncated TβRII mRNA transcript (fig 1). TGF-β2 mRNA was below the level of detection in the pancreas of both groups of mice, even after caerulein administration (data not shown).
Serum amylase, lipase and TAP levels
Acute pancreatitis is associated with increases in serum amylase and lipase levels. Therefore, we sought to compare serum levels of these digestive enzymes in FVB−/− and FVB+/−mice (fig 2). In FVB−/− mice receiving sham injections, mean serum values for amylase and lipase were 2120 (240) U/ml and 104 (28) U/ml, respectively. Caerulein injections in these mice were associated with a sixfold increase in serum amylase levels and a 36-fold increase in serum lipase levels, and these increases were highly significant (p<0.002). In FVB+/− mice, basal serum levels were 2420 (76) U/ml for amylase and 161 (23) U/ml for lipase. Caerulein treatment caused very slight increases in serum amylase (64%) and lipase (39%) levels in FVB+/− mice.

Serum amylase and lipase levels. Amylase (A) and lipase (B) levels were measured in serum samples from wild-type (FVB−/−) or transgenic (FVB+/−) mice that were either injected with buffer (dark grey bars) or with caerulein (light grey bars), as indicated in the Methods section. Values are means (SEM) of six animals per group. *p<0.002 compared with respective sham controls.
Because of the slight increases in serum lipase levels following caerulein injections in the FVB+/− mice, we next examined serum TAP levels, taken as a measure of the extent of pancreatic trypsin activation. Serum TAP levels increased 3.3-fold in FVB−/− mice after caerulein administration (fig 3). In contrast, there was only a slight increase in serum TAP levels in FVB+/− mice, and this increase was not statistically significant (p = 0.2).
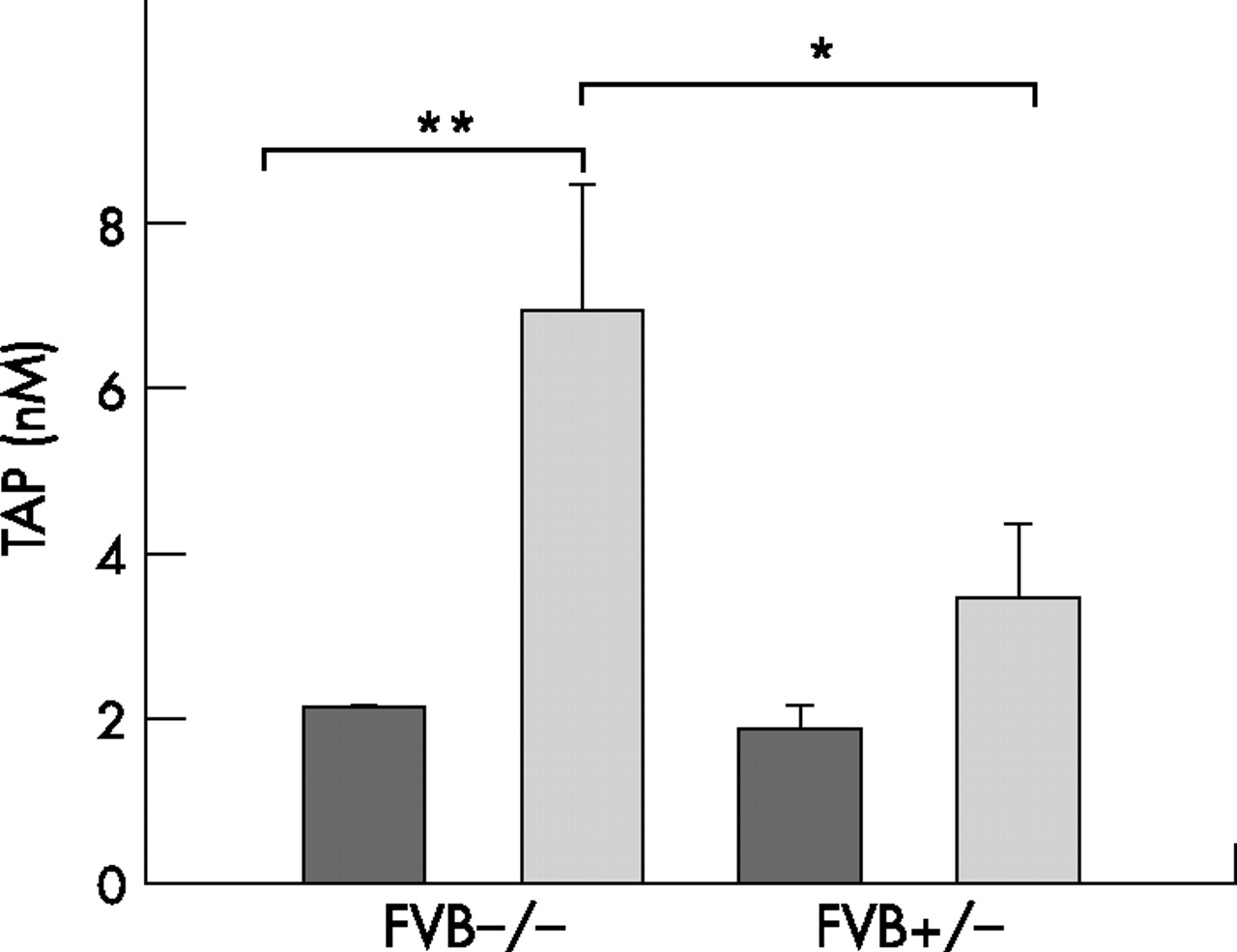
Serum trypsinogen activation peptide (TAP) levels. TAP levels from wild-type (FVB−/−) or transgenic (FVB+/−) mice that were injected with either buffer (dark grey bars) or caerulein (light grey bars) were measured as indicated in the Methods section. *p<0.012 compared with caerulein treated FVB−/− mice; **p<0.002 compared with the corresponding control. Values are means (SEM) of six animals per group.
Evaluation of pancreatic oedema
There was a marked difference in the severity of pancreatitis between wild-type and transgenic mice. In FVB−/− mice, sham injections did not grossly alter the appearance of the pancreas (fig 4A) whereas caerulein administration was associated with an enlarged pancreas that was grossly wet and showed prominent vessels suggestive of an acute inflammatory process (fig 4B). In FVB+/− mice, the pancreas was slightly smaller (fig 4C) than the pancreas of the wild-type control group, and caerulein administration was associated with only a slight increase in pancreatic size, without grossly visible changes suggesting either oedema or inflammation (fig 4D).

Gross morphology. Pancreata from wild-type (A–B) or transgenic (C–D) mice were removed 16 h after the last injection of buffer (A, C) or caerulein (B, D). The scale shown is in centimetres.
To determine the percentage of pancreatic water in the various groups, tissues were dried for 36 h at 90°C. The resulting dry weights were subtracted from the original wet weights, yielding an estimate of the water content of the pancreata.20 The relative water content was calculated by dividing this value by the wet weight and multiplying by 100.20 In the pancreas of FBV−/− mice (n = 5), caerulein injections significantly (p<0.0002) increased this value from 71.5% to 82% (fig 5). In contrast, the pancreata of FBV+/− mice (n = 6) exhibited only slight increases in the relative water content following caerulein injections, from 70.8% to 71.4% (p = 0.51).

Pancreatic wet weight. Wet weight was determined in pancreata from wild-type (FVB−/−) or transgenic (FVB+/−) mice that were injected with either buffer (dark grey bars) or caerulein (light grey bars), as indicated in the Methods section. Values are means (SEM) of six animals per group. *p<0.002 compared with respective sham controls.
Histological analysis comparing FBV−/− and FBV+/− mice
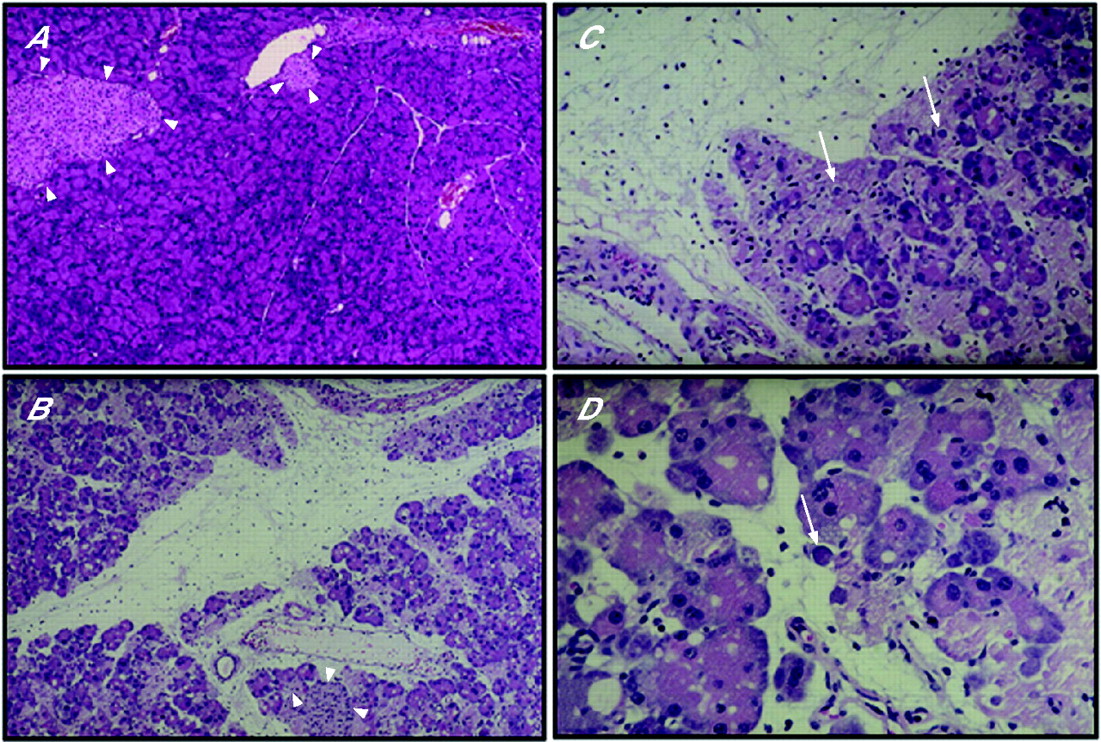
As expected, none of the six pancreatic samples from wild-type control mice harboured any inflammatory cells (fig 6A), and the endocrine islets appeared normal (arrowheads). In the caerulein treated group, three pancreatic samples exhibited marked oedema (fig 6B) and all six samples exhibited a typical pattern of inflammatory cells, consisting mainly of neutrophils and/or macrophages (fig 6B–D), in association with the presence of apoptotic bodies (fig 6C, D) but with normal appearing islets (arrowheads, fig 6B). In addition, apoptotic bodies were visible throughout the pancreas (arrows, fig 6C, D). In contrast, four of six pancreatic samples from transgenic control mice exhibited mild and variable degrees of macrophage and lymphocytic infiltration and occasional apoptotic bodies (fig 7A). This increased lymphocytic/macrophage infiltration has been attributed to the increased levels of TGF-β1 that is observed in the pancreas in this model.12 Following treatment with caerulein, none of the six pancreata from the transgenic mice exhibited oedema or acinar cell degeneration (fig 7B–D). Instead, there was only a mild increase in the number of neutrophils and/or macrophages, and an occasional appearance of apoptotic bodies (fig 7C, D). The results of histological scoring are summarised in table 2.
Scoring of the histological specimens by morphological criteria

Histological analysis in wild-type mice. Pancreatic sections (4 μM thick) from control mice that were injected with buffer (A) or caerulein (B–D) were stained with haematoxylin-eosin. Normal pancreatic architecture is evident in sham injected control mice, including normal endocrine islets that are indicated by arrowheads (A). Caerulein injection resulted in marked oedema without altering the islets (B, arrowheads) but with the appearance of strong leucocytic infiltration (B–D). In addition, there is disruption of the acinar cell architecture and the appearance of apoptotic bodies, which are indicated by arrows (C). An apoptotic body (spherical eosinophilic structure) is readily evident in D (arrow), and the presence of polymorphonuclear cells is also evident (D). Magnification: A, 100×; B, 180×; C, 200×; D, 250×.

Histological analysis in transgenic mice. Pancreatic sections (4 μM thick) from FVB+/− mice that were injected with buffer (A) or caerulein (B–D) were stained with haematoxylin-eosin. Foci of lymphocytic/macrophage infiltrates are present in the pancreas of sham injected mice (A). Following injection of caerulein, the pancreatic parenchyma remains compact, indicating the absence of oedema, and acinar cell disorganisation is not observed (B–D). There is a slightly increased infiltration by lymphocytes, and only rare polymorphonuclear cells (arrow) are present (D). Apoptotic bodies are seen in (C) and (D) (indicated by arrowheads). Magnification: A, 100×; B, 180×; C, D, 250×.
Amylase secretion studies
The above in vivo results do not indicate whether there is a defect in the acinar cell of TβR+/− mice that causes resistance to caerulein or whether the defect is due to alterations either in other cellular components in the pancreas or with respect to caerulein pharmacology in the transgenic mice. Because early events in the acinar cell that are known to occur in caerulein induced acute pancreatitis can be recapitulated in vitro by exposing pancreatic acini to supramaximal concentrations of the secretagogue,23–26 we next determined whether pancreatic acini from transgenic mice are resistant to CCK receptor activation in vitro. In this series of studies we used CCK8, the C terminal octapeptide of CCK, which binds and activates the acinar cell CCK receptor with the same high affinity and to the same extent as caerulein.
In acini isolated from normal FBV−/− mice, CCK8 exerted a classical biphasic effect on amylase release (fig 8). Maximal stimulation occurred at 0.1 nM CCK8 whereas higher concentrations of CCK8 exerted a progressively smaller effect on secretion. This phenomenon of restricted stimulation by supraphysiological concentrations CCK8 has been recognised for three decades and corresponds to the concentrations of ligand that induce acute pancreatitis-like changes in the acinar cell.23–27 It has been attributed to several mechanisms, including disorganisation of microtubules, membrane blebbing and excessively sustained rise in intracellular free calcium levels.23–29

Effects of the C terminal octapeptide of cholecystokinin (CCK8) on amylase release. Acini isolated from FVB−/− mice (•) and from FVB+/− mice (▪) were incubated for 30 min in the absence or presence of the indicated CCK8 concentrations, and amylase levels in the medium were determined. Data are expressed as per cent increase in amylase release with respect to control values, and are means (SEM) of duplicate determinations from three separate experiments per group.
In acini isolated from FBV+/− mice there was a markedly altered pattern of amylase secretion. Thus maximal stimulation occurred at a 100-fold greater concentration of CCK8 than in FBV−/− mice (1 nM), and even higher concentrations of CCK8 failed to exhibit a marked attenuation of secretion (fig 8). Furthermore, at the respective maximally effective concentrations of CCK8, the amount of amylase released as a per cent of total amylase content was greatly attenuated in acini from the FBV+/− mice. Thus at its maximally effective concentration in acini from FBV−/− mice (0.1 nM) or FBV+/− mice (1 nM), the per cent of total amylase content that was released by CCK8 was 18 (SD1) and 8.5 (SD0.3), respectively. By contrast, LDH levels were comparably low in the supernatant of acini from both groups of mice, indicating that acini from the transgenic mice were not exhibiting enhanced cellular damage.
Characterisation of CCK receptor expression in pancreatic acini
The failure of caerulein to induce typical features of acute pancreatitis in FBV+/− mice and the markedly altered secretory response to CCK8 in acini isolated from these mice raised the possibility that CCK receptor expression was altered in acini from these transgenes. Therefore, pancreatic CCK receptor mRNA expression was characterised next. The CCK A receptor (CCK-AR) mRNA transcript (2.7 kb) was present in RNA samples isolated from the pancreas of FBV−/− sham (fig 9). There was a reproducible decrease in the steady state levels of pancreatic CCK-AR mRNA following caerulein injection (fig 8). In contrast, the pancreas of FBV+/− mice exhibited high CCK-AR mRNA levels, both before and after administration of caerulein (fig 9). The CCK receptor B (CCK-BR) mRNA transcript was below the level of detection in both groups.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of cholecystokinin A receptor (CCK-AR). Total pancreatic RNA (20 μg/lane) was isolated from wild-type (FVB−/−) and transgenic (FVB+/−) mice that were injected with either buffer (−) or with caerulein, as described in the Methods section. Northern blot analysis was then carried out using a 32P-labelled cDNA probe (500 000 cpm/ml) specific for CCK-AR. A 7S ribosomal cDNA probe (50 000 cpm/ml) was used as a loading and transfer control. Exposure times were overnight for CCK-AR and 3 h for 7S.
DISCUSSION
Caerulein induced acute pancreatitis is a commonly used animal model to investigate the pathophysiology of this disorder. Caerulein injections cause the premature activation of zymogen enzymes in pancreatic acinar cells which is believed to be a critical step in the initiation of acute pancreatitis.4,30–32 In the present study we documented that transgenic mice that bear a dominant negative type II TGF-β receptor, which is predominantly expressed in the pancreas and liver in this model,12 failed to develop most of the characteristics of acute pancreatitis. After repeated injections of caerulein, wild-type mice showed mild to moderate oedema in the pancreas, as determined by gross morphology, and an increase in pancreatic water content. In contrast, the pancreas of transgenic mice that failed to develop pancreatic oedema following caerulein administration did not exhibit an increase in pancreatic water content.
Two previous studies in which control mice and mice expressing either a dominant negative form of TβRII that interferes with TGF-β signalling,33 or a soluble form of TβRII that sequesters TGF-β ligands,34 were injected with caerulein for 3 to 10 weeks to induce recurrent episodes of pancreatitis, and the effects on pancreatic fibrosis were evaluated. Fibrosis was markedly attenuated in the pancreas of both mouse models. However, neither study examined the short term effects of caerulein in their models. Thus neither study considered the possibility that the decreased fibrosis could have been due to attenuated induction of acute pancreatitis in these models.
Several lines of evidence in the present study suggest that this lack of oedema was a result of a marked reduction in the capacity of caerulein to induce acinar cell injury in transgenic mice. First, serum lipase levels, which are sensitive indicators of acute pancreatic cell damage, were not elevated and serum amylase levels were only slightly elevated in transgenic mice following caerulein administration. In contrast, wild-type mice exhibited a 36-fold increase in serum lipase levels after caerulein treatment. Although there was a slight increase in serum amylase levels following caerulein administration to the transgenic mice, this increase could be due to stress associated stimulation of parotid amylase release.35 Second, caerulein induced a marked and significant increase in serum TAP levels in wild-type mice. TAP corresponds to the N-terminal 7–10 amino acids that are released following the activation of trypsinogen into active trypsin, and the presence of high TAP levels in the serum of these mice reflects inappropriate and premature activation of trypsinogen.29 By contrast, there was only a slight increase in serum TAP levels in the FBV+/− mice, underscoring the inability of caerulein to induce acute pancreatitis in these transgenes. Third, caerulein administration in the transgenes expressing the truncated TβRII caused only a slight increase in TGF-β1 mRNA levels and did not alter TGF-β3 mRNA levels. This is in sharp contrast with the marked rise in TGF-β1 mRNA levels induced by caerulein in control mice. Taken together with the absence of gross oedema and the pancreatic wet weight results, these observations suggest that expression of a truncated TβRII modulates the ability of caerulein to activate the pathways that lead to acute pancreatitis.
Histological examination of the pancreas from wild-type mice with pancreatitis revealed acinar cell necrosis and typical infiltration of inflammatory cells into the parenchyma of the pancreas. These infiltrates mainly consisted of neutrophils and macrophages. In contrast, caerulein administration did not result in acinar cell necrosis in the transgenes. Furthermore, the predominant inflammatory cells in the pancreas of these mice consisted of lymphocytes and macrophages. Previous studies have demonstrated that neutrophils express TGF-β receptors, and that TGF-β acts to induce their migration and to potentiate the effects of inflammatory cytokines on their migration and accumulation at sites of inflammation.36–38 The paucity of neutrophils in the pancreas of the transgenes could conceivably be due to the failure of caerulein to induce acinar cell damage as well as to an abrogation by the truncated TβRII of the mechanisms that activate neutrophil recruitment. Furthermore, in the rat, neutrophil depletion prior to caerulein treatment results in a decrease in acinar cell necrosis and an increase in apoptotic acini.11 It is possible, therefore, that the absence of acinar cell necrosis in the pancreas of the transgenes may be due, at least in part, to the smaller number of neutrophils in these mice.
The in vitro secretion results in the present study indicate that expression of a truncated TβRII modulates the effects of caerulein on the acinar cell secretory apparatus. This was evidenced by a shift to the right in the secretory dose–response curves in transgene derived acini, by the failure to achieve a maximal stimulatory effect on secretion that is comparable with that observed in normal mice, and by complete disappearance of the phenomenon of restricted stimulation of secretion at high CCK8 concentrations. Moreover, these alterations were not due to a generalised defect in the actions of calcium metabolising pancreatic secretagogues, inasmuch as the secretory actions of bombesin, which acts through its own receptor, were not altered in these acini (unpublished observations). These observations suggest that there may be selective resistance to the actions of CCK8/caerulein when TGF-β signalling is attenuated in the pancreatic acinar cell.
The failure of caerulein to induce acute pancreatitis in the FBV+/− mice and the altered actions of CCK8 in acini from these mice raised the possibility that these transgenes express fewer CCK receptors. However, northern blotting revealed that the pancreata of FBV+/− mice exhibited higher levels of the CCK-AR mRNA transcript than the pancreata of FBV−/− mice. Although the reasons for the increased expression of CCK-AR in these transgenes are not readily evident, their pancreas exhibits enhanced acinar cell proliferation and increased apoptosis.12 It is possible therefore that the pancreas in these transgenes contains a younger population of acini that express higher levels of CCK-AR. In support of this hypothesis, increased CCK-AR expression has been demonstrated following pancreatic regeneration.39 By contrast, neither group expressed CCK-BR, indicating that the altered responses to caerulein and CCK8 were not due to alterations in the expression of CCK receptor subtypes. The fact that caerulein decreased CCK-AR mRNA levels in wild-type but not in the FBV+/− mice raises the possibility that proper TGF-β signalling may be required for modulation of CCK-AR mRNA expression.
A number of studies using transgenic mice have provided new insight into the pathophysiology of acute pancreatitis. Thus the important role of oxidative stress in acute pancreatitis progression was highlighted by the observation that transgenes overexpressing copper–zinc superoxide dismutase exhibit an attenuated form of acute pancreatitis following repeated injections of caerulein.40 Similarly, the contribution of the inflammatory process in the pancreas during acute pancreatitis progression was underscored by the observation that mice lacking the neurokinin 1 receptor, which is activated by the proinflammatory cytokine substance P, exhibit less severe pancreatitis in the caerulein model.41 The fact that cathepsin B is an important intracellular activator of intracellular trypsinogen was demonstrated using cathepsin B knockout mice, which also exhibited less severe acute pancreatitis following repeated administration of caerulein.42 In a similar vein, the present findings suggest that a functional TGF-β signalling pathway is essential for the induction of acute pancreatitis by caerulein, raising the possibility that there is crosstalk between TβRII and CCK-AR signalling pathways.
In a previous study of the potential role of TGF-β in acute pancreatitis, a dominant negative mutant form of TβRII was expressed in the pancreas under control of the pS2 (TFF1) mouse trefoil peptide promoter.43 In this model, the mice exhibited increased susceptibility to caerulein induced pancreatitis, as evidenced by the presence of severe pancreatic oedema and inflammatory cell infiltration. This excessive responsiveness to caerulein was associated with enhanced activation of T- and B-cells, in conjunction with IgG-type autoantibodies against the pancreatic acinar cell and IgM-type autoantibodies against pancreatic ductal cells. The authors concluded that TGF-β signalling either suppresses autoimmunity or maintains the integrity of the pancreatic acinar cell. It should be noted, however, that immunocytes present in the thymus, spleen, bone marrow and lymph nodes have been found to express and synthesise trefoil proteins,44 raising the possibility that these cell types may have a role in immune regulation that was perturbed by the expression of dominant negative TβRII.
To date, interest in the role of TGF-β in acute pancreatitis has focused mainly on pancreatic repair and fibrosis. Thus pancreatic TGF-β expression is increased in the first hours after induction of acute pancreatitis in rats9 and in patients with chronic pancreatitis.45 These observations suggest that TGF-βs may exert a deleterious effect in the fibrotic pancreas. In humans, TGF-β1 expression is increased in the pancreas of patients suffering from acute necrotising pancreatitis.46 Moreover, TGF-β1 levels are increased in the serum of patients with acute pancreatitis.47 Together with our current findings, these observations raise the possibility that TGF-βs may have a role in the pathogenesis of acute pancreatitis in humans. It will be important, therefore, to determine whether pharmacological interventions that abrogate TGF-β signalling, such as small molecule inhibitors that interfere with TGF-β receptor serine–threonine kinase activity and the downstream signalling events,48 may ultimately have a role in the treatment or prevention of acute pancreatitis.
REFERENCES
Footnotes
-
Published Online First 28 November 2006
-
This work was supported by Public Health Service Grant CA-75059 awarded by the National Cancer Institute to MK. SW was the recipient of postdoctoral trainee awards from the Ursula Zindel-Hilti-Foundation, the Ciba-Geigy-Anniversary Foundation and the Department of Visceral and Transplantation Surgery of the University Hospital of Bern.
-
Competing interests: None.
-
Current addresses: *Department of Visceral and Transplantation Surgery, University of Zurich, Zurich, Switzerland; †Department of Gastroenterology, Endocrinology and Nutrition, Ernst-Moritz-Arndt University of Greifswald, Greifswald, Germany