Article Text

Abstract
Objectives: The intestinal mucosa is constantly exposed to a dense and highly dynamic microbial flora and challenged by a variety of enteropathogenic bacteria. Antibacterial protection is provided in part by Paneth cell-derived antibacterial peptides such as the α-defensins. The mechanism of peptide-mediated antibacterial control and its functional importance for gut homeostasis has recently been appreciated in patients with Crohn’s ileitis. In the present study, the spatial distribution of antimicrobial peptides was analysed within the small intestinal anatomical compartments such as the intestinal crypts, the overlaying mucus and the luminal content.
Methods: Preparations from the different intestinal locations as well as whole mouse small intestine were extracted and separated by reversed-phase high-performance liquid chromatography. Antibacterial activity was determined in extracts, and the presence of antimicrobial peptides/proteins was confirmed by N-terminal sequencing, mass spectrometry analysis and immunodetection.
Results: The secreted antibacterial activity was largely confined to the layer of mucus, whereas only minute amounts of activity were noted in the luminal content. The extractable activity originating from either crypt/mucus/lumen compartments respectively (given as a percentage) was for Listeria monocytogenes, 48 (4)/44 (4)/8 (8); Enterococcus faecalis, 44 (10)/49 (3)/7 (7); Bacterium megaterium, 56 (4)/42 (3)/2 (1); Streptococcus pyogenes, 48 (4)/46 (3)/6 (6); Escherichia coli, 46 (4)/47 (3)/7 (7); and Salmonella enterica sv. Typhimurium, 38 (3)/43 (7)/19 (10). A spectrum of antimicrobial peptides was identified in isolated mucus, which exhibited strong and contact-dependent antibacterial activity against both commensal and pathogenic bacteria.
Conclusion: These findings show that secreted antimicrobial peptides are retained by the surface-overlaying mucus and thereby provide a combined physical and antibacterial barrier to prevent bacterial attachment and invasion. This distribution facilitates high local peptide concentration on vulnerable mucosal surfaces, while still allowing the presence of an enteric microbiota.
Statistics from Altmetric.com
Production of antimicrobial peptides is one of the major strategies used by multicellular organisms to control microbial growth. It allows efficient restriction of the normal microflora without induction of a potentially harmful inflammatory response and protects against pathogens.1 Paneth cells, situated at the base of the small intestinal crypts, are important contributors to the small intestinal antimicrobial barrier through synthesis and release of antimicrobial peptides and proteins such as enteric α-defensins (in mice also called cryptdins), CRS (cryptdin-related sequences) peptides, lysozyme and secretory phospholipase A2.2–6 Cholinergic stimuli or bacterial compounds have been suggested to induce secretion.7 8 Bacterial colonisation is, however, not a prerequisite for synthesis of α-defensins since the peptides are also produced in germ-free mice.9 Like other peptide antibiotics, defensins and CRS peptides are produced as preproforms that are further processed to yield the mature active bactericidal peptide.2 5 9
The functional importance of α-defensins in antibacterial protection was shown in knock-out mice lacking the processing enzyme matrilysin (MMP7) and hence active peptides.10 In contrast, mice engineered to produce an additional human defensin in the gut were less sensitive to Salmonella infection.11 In humans, reduced levels of α-defensins were found in patients with Crohn’s disease,12 13 suggesting a role for enteric α-defensins in the pathogenesis of inflammatory bowel disease. It is noteworthy that Crohn’s patients displayed a thinner layer of mucus and were colonised with pathogenic bacteria.14 We have recently shown that patients exposed to orally acquired microorganisms after gastric bypass surgery have increased secretion and expression of α-defensins in the small intestine, indicating the importance of antimicrobial peptides in the modulation of intestinal microbial.15
In the present study, we biochemically characterise enteric antimicrobial peptides and proteins of the intestine and examine their anatomical distribution in small intestinal compartments. The antimicrobial peptides and proteins were retained in the mucus layer covering the intestinal epithelium and revealed contact-dependent antibacterial activity against commensal and pathogenic bacteria. The results suggest that the peptide-containing mucus layer can significantly contribute to limit bacterial growth to the intestinal lumen and protect the integrity of the intestinal epithelium.
MATERIALS AND METHODS
Preparation of small intestine, primary crypts and mucus
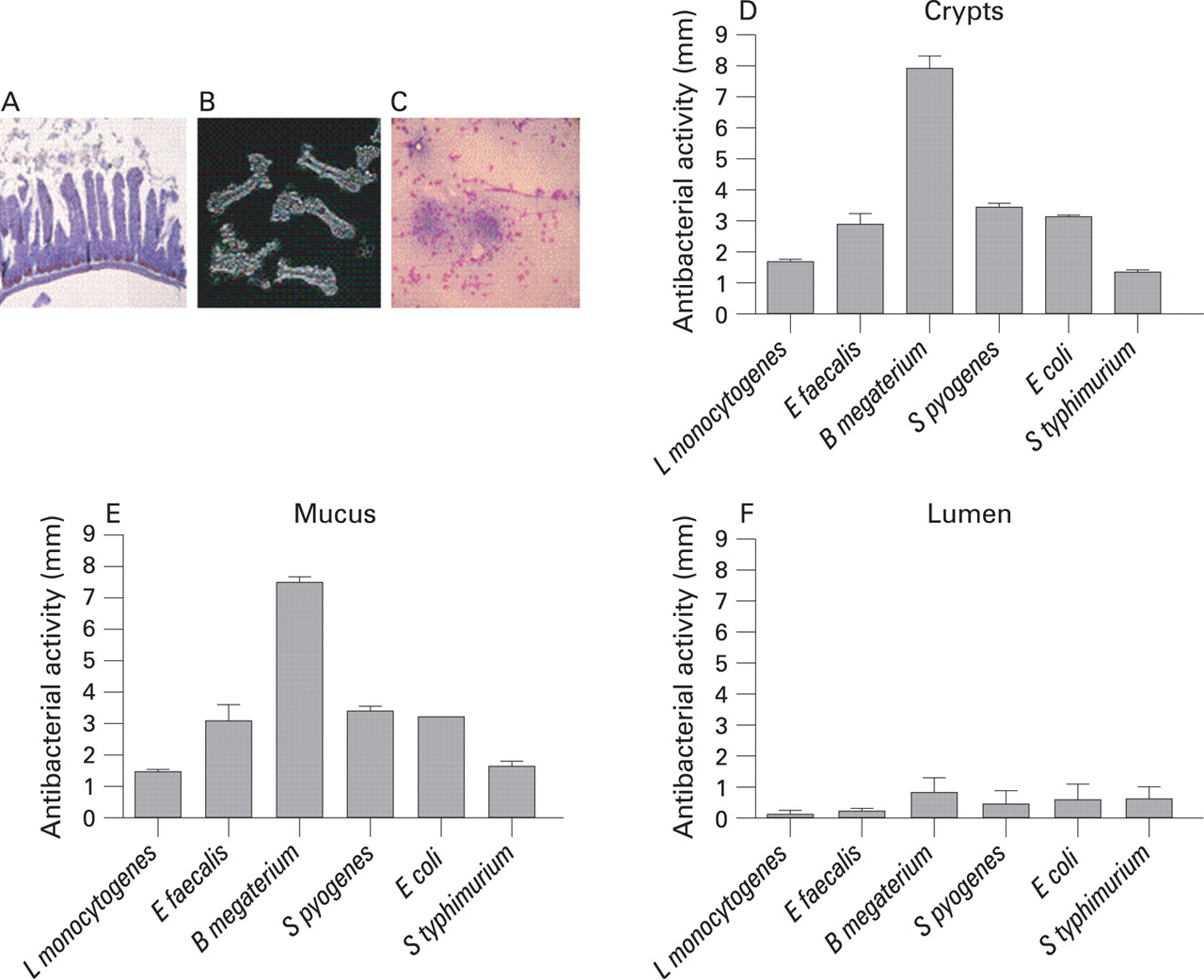
C3H/HeN freely feeding mice, 6–8 weeks old, were kept under specific pathogen-free (SPF) conditions approved by the local animal legislation. Mice were euthanised and the small intestine was removed. Intestinal luminal content was obtained by gently flushing the intestine (attached to a plastic syringe) with 10 ml of ice-cold phosphate-buffered saline (PBS). The resulting liquid was concentrated using lyophilisation. Mucus was collected from PBS-washed and longitudinally opened intestine by mild physical separation using round forceps. Crypts were prepared from inverted tissue by incubation in 30 mM EDTA at 37°C for 10 min, followed by mild agitation and differential centrifugation, as described.16 Small intestine was fixed in formalin and stained with H&E for morphology and with antilysozyme (Dako, Glostrup, Denmark). Isolated crypts were inspected under light microscopy. Mucus was smeared out on a glass slide, air-dried, heat fixed and stained with Kwik-Diff stain (Shandon, Pittsburgh, Pennsylvania, USA). For whole-organ extraction, small intestine, from each mouse, was ground in liquid nitrogen to obtain a fine powder and subsequently extracted for 3 h (repeated vortexing for 2 min) with ice-cold 60% aqueous acetonitrile containing 1% trifluoroacetic acid (TFA), final concentrations.9 Crypts, lumen and mucus preparations were sonicated at 100 W for 20 s on ice, prior to extraction. The extracts were centrifuged at 16 000 g for 20 min and the obtained supernatants were freeze-dried, re-dissolved in 0.5 ml of water with 10% ethanol, and cleared by centrifugation. Protein concentrations were measured using DC Protein Assay reagent (Bio-Rad Laboratories, Hercules, California, USA) according to the manufacturer. The protein concentrations of the mucus, crypt and small intestinal extracts were approximately 8, 3.3 and 4.5 mg/ml, respectively, and the luminal content extracts were approximately 0.6 mg/ml.
High-performance liquid chromatography (HPLC) purification and analysis of purified components
Components in extracts were separated by reverse phase (RP)-HPLC as described before.9 Extracted material representing one-third of an intestine was applied from each preparation. In brief, elution was performed with 0.18% TFA in H2O (solvent A) and a gradient of 0.15% TFA in acetonitrile (solvent B) on a 0.46×25 cm Vydac C18 column (The Separation Group, Hesperia, California, USA). Fractions of 0.8 ml were collected and lyophilised. Freeze-dried chromatographic fractions were re-dissolved in 10 µl of 10% ethanol. N-terminal sequence analysis was done by Edman degradation with a Procise cLC or HT instrument (PE Applied Biosystems, Foster City, California, USA). Matrix-assisted laser desorption-ionisation time-of-flight mass spectrometry (MALDI-TOF MS) analysis was carried out with a Reflex III (Bruker Daltonics, Leipzig, Germany).9 For immunoblotting, samples were mixed with NuPAGE LDS Sample buffer (Invitrogen, San Diego, California, USA), adjusted to 10% β-mercaptoethanol and heated before loading. Proteins were separated on a 1.0 mm 10% or 4–12% NuPAGE Bis-Tris gel (Invitrogen) in NuPAGE MES SDS running buffer (Invitrogen) and then blotted onto polyvinylidene difluoride (PVDF) filters (Invitrogen). The filters were blocked for 1 h in PBS containing 1% Tween (PBST) supplemented with ECL advance blocking reagent (Amersham Pharmacia Biotech, Uppsala, Sweden). Incubation with antigen-specific antibodies or horseradish peroxidase (HRP)-conjugated detection antibodies was for 1 h each at room temperature in PBST. Antiserum against the N-terminal part of ribosomal protein L29 was raised in rabbits (Innovagen, Lund, Sweden); antilysozyme P was purchased from Calbiochem (San Diego, California, USA). Anti-MBD-1 and anti-mBD-3 were from Santa Cruz Biotechnology (Santa Cruz, California, USA). Antibody reactivity was detected by chemiluminescence with SuperSignal West Dura (Pierce Biotechnology, Rockford, Illinois, USA) or alternatively ECL Advance, and evaluated by densitometry using Intelligent DarkBoxII (FujiFilm, Tokyo, Japan).
Determination of antibacterial activity
Bacterial strains used were Escherichia coli strain D21, Salmonella enterica sv. Typhimurium ATCC 14028, Listeria monocytogenes type 1 clinical isolate, Bacillus megaterium strain Bm11, Enterococcus faecalis ATCC 29212 and a clinical isolate of Streptococcus pyogenes. A low-salt agarose thin layer assay was used as described17 but with an overnight incubation at 30°C, after which the bacterial growth inhibition was recorded. A dilution series of extracts from each individual (crypt, mucus, lumen) was analysed on the same plate and served as vehicle control. Resistin-like molecule β (RELMβ) was from Pepro Tech Inc. (Rocky Hill, New Jersey, USA). Activity is presented as the diameter of the inhibition zone minus the diameter of the sample well (3 mm). The inhibition zone was converted to activity units and the distribution of segmental activity was calculated and presented as mean (SEM).18 Dilution series were analysed by two-way analysis of variance (ANOVA; Prism, GraphPad Software Inc., San Diego, California, USA).
Mucus-associated antibacterial activity
Bacillus megaterium, L monocytogenes, E faecalis, E coli and Shigella flexneri dysenteriae were grown overnight on blood agar plates, resuspended in PBS, and diluted to an optical density of 0.5 at λ = OD600. For functional analysis, mucus was isolated from C3H/HeN mice, diluted 1:3 in PBS, and suspended thoroughly. Resuspended mucus or PBS (150 µl per well in 96-well plates) was centrifuged at 2500 rpm at 4°C for 10 min and the liquid supernatant was removed. Bacillus megaterium was diluted in PBS and 200 µl per well was added (106 bacteria) and the plates were either left untreated (liquid bacteria) or centrifuged at 1500 rpm for 15 min (mucus-associated bacteria). Subsequently, the samples were resuspended by repeated pipetting and plated on LA plates in serial dilution to obtain the number of viable bacteria per well. Experiments were performed in triplicate. Results were analysed by t test.
To visualise mucus-associated killing, 150 µl of mucus per well was centrifuged on 8-well chamber slides and the liquid supernatant was removed. As a control, matrigel gel (BD Biosciences, Franklin Lakes, NJ, USA) was carefully thawed on ice, placed 90 µl per well on a precooled 8-well chamber slide and incubated at 37°C to solidify the gelatinous matrix. Bacteria (4×106 in 200 µl) were added and the slide was centrifuged at 1500 rpm for 15 min at room temperature. Live bacteria were distinguished from dead by the LIVE/DEAD BacLight bacterial viability kit (Molecular Probes, Eugene, Oregon, USA). A 3 µl aliquot of solution A and B was diluted in 800 µl of PBS and added to the wells. The slides were incubated in the dark for 15 min before the liquid supernatant was removed and the slides were covered with a coverslip. Green (live) or red (dead) bacteria were immediately visualised using an Apotome (Zeiss, Jena, Germany) at a magnification of ×630.
RESULTS
Antimicrobial activity against commensal and enteropathogenic bacteria in protein extract of different compartments of the intestine
To determine the spatial distribution of antibacterial peptides within the small intestine, we prepared and collected isolated crypts, mucus as well as luminal content from wild-type mice. The cellular composition of different preparation prior to chemical extraction is illustrated in fig 1A–C. Equal volumes of extracts, representing identical starting segments of the small intestine, were sequentially diluted and applied to thin agar plates with growing bacteria. High antibacterial activity was recorded with extract from crypt and mucus against all bacteria tested except S typhimurium (fig 1D,E). The extractable activity originating from either crypt/mucus/lumen compartments, respectively (given as a percentage of the sum of activity from crypt, mucus and lumen against different bacteria) was: L monocytogenes, 48 (4)/44 (4)/8 (8); E faecalis, 44 (10)/49 (3)/7 (7); B megaterium, 56 (4)/42 (3)/2 (1); S pyogenes, 48 (4)/46 (3)/6 (6); E coli, 46 (4)/47 (3)/7 (7); and S Typhimurium, 38 (3)/43 (7)/19 (10). The luminal wash exhibited significantly (p<0.01) lower activity than crypt and mucus against L monocytogenes, E faecalis, B megaterium, S pyogenes and E coli, whereas the lower activity against S Typhimurium was not significant (fig 1F). For comparison, the unrelated recombinant small intestinal protein RELMβ did not display any activity at a concentration of 2 mg/ml (data not shown).
Detailed biochemical analysis of isolated proteins
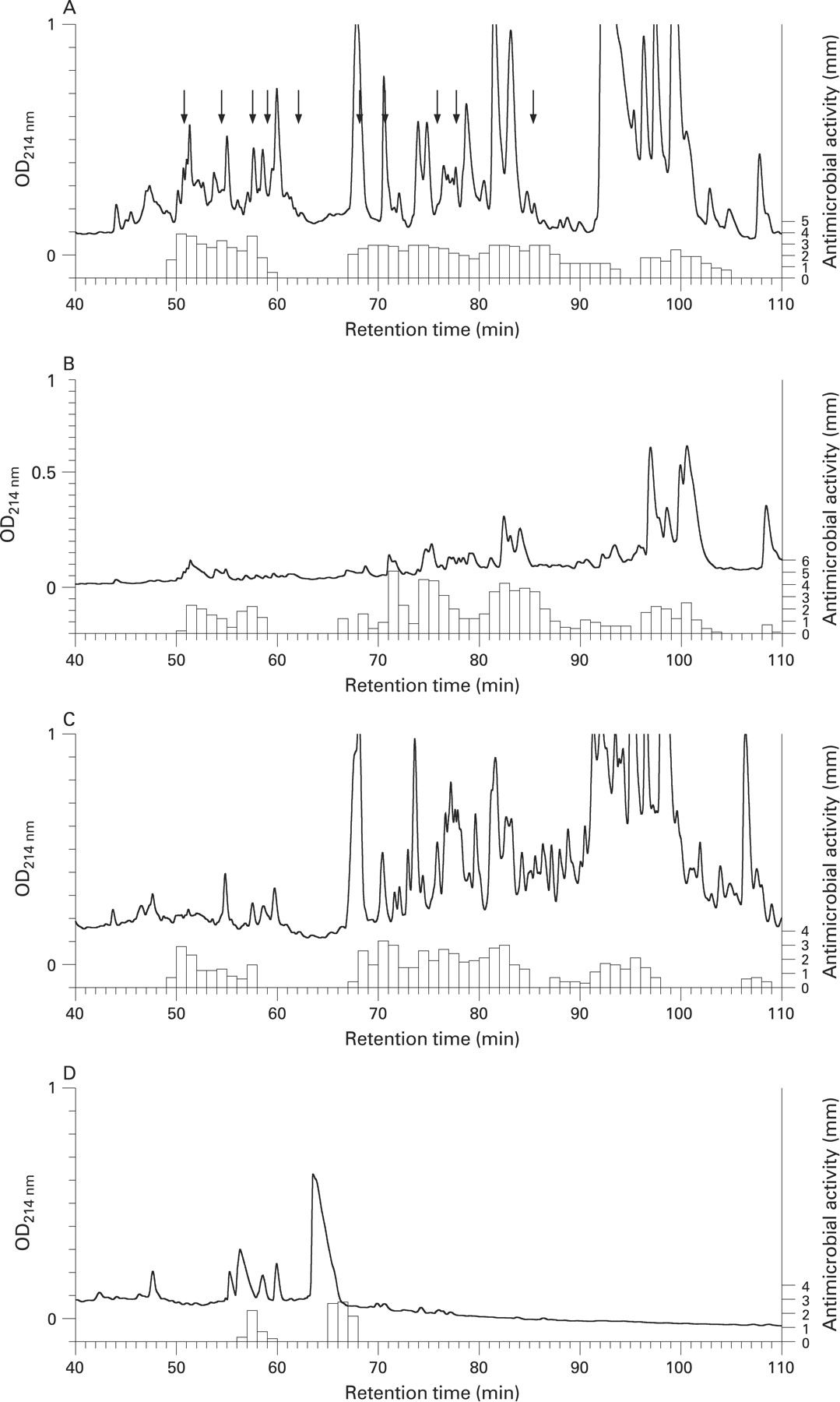
Extracts from different small intestinal compartments were fractionated by RP-HPLC and the resulting fractions were assayed for antibacterial activity against S pyogenes using a thin layer assay (fig 2A–D). The eluting profile of antibacterial activity in total small intestine and isolated crypts was similar (fig 2A,B) due to the fact that the majority of antibacterial activity in the intestinal tract is produced by Paneth cells. Additionally, a similar elution pattern of antibacterial activity was found in mucus preparations, whereas only minor activity was noted in the luminal wash (fig 2C,D). We next focused on identifying known Paneth cell constituents (eg, α-defensins, CRS4C peptides, lyzozyme P and phospholipase A2) and establishing the identity of the antibacterial compounds in other fractions with notably high antibacterial activity. α-Defensins and CRS4C peptides eluted in a region corresponding to fractions 50–62 (table 1). Ubiquicidin, which possesses antimicrobial activity and is identical to ribosomal protein S30,19 eluted in fraction 57. α-Defensin 1–4, 6, CRS4C peptides and ubiquicidin were detected in both total small intestine and isolated crypts, again reflecting the similar antibacterial composition in these preparations (fig 2A,B, table 1).

The high antibacterial activity noted in mucus extract (fig 1E) suggested that components released from the epithelial cell layer might end up in the mucus layer. Indeed, the mucus extract separated by RP-HPLC presented an elution profile of antibacterial activity with strong similarities to that of both small intestine and isolated crypts. In the first region (fig 2C fractions 50–60) we could identify α-defensin 1–4 and 6, as well as ubiquicidin, with MALDI-TOF (fig 3A–F).

Since β-defensins are believed to play a role in inflammatory bowel diseases,28 we screened particularly for the expression of the mouse β-defensins mBD-1 and mBD-3 using immunoblot techniques. Expression of mBD-1 and mBD-3 could, however, not be detected in the extracts of the total intestinal tissue (data not shown).
A second region of components with antibacterial activity was found in fractions 68–90 eluting from small intestine, crypt and mucus. Four ribosomal proteins, L29, L26, L38 and L35, were detected in different fractions of total small intestinal extract (table 1). These proteins were identified by their N-terminal primary structure and in most cases confirmed by MALDI-TOF MS (table 1). Additionally, this region contained two important antibacterial enzymes, phospholipase A2-IIA and lysozyme P. Lysozyme P could be identified by immunoblot in the mucus but at lower levels than in small intestine or crypt material (data not shown). Accordingly, the antibacterial activity in the fractions containing lysozyme (fraction 86) was lower in mucus than in small intestine/crypts (fig 2).
A third region with activity was found in fractions 95–105 in both total small intestinal tissue and isolated crypts, and in fractions 90–100 in mucus (fig 2). We have not been able to identify components in this region primarily because of difficulties in separating the active component from the bulk of other components eluting in this region. The luminal extract had few regions of significant activity (fig 2D). The large number of fractions presenting antibacterial activity is a consequence of extracting a tissue with multiple active compounds, using a sensitive bacterium and an assay that measures a combination of bactericidal and bacteriostatic activity.
Collectively this shows that components produced by Paneth cells as well as products most probably originating from the other intestinal epithelial cells can be found as active antimicrobial peptides/proteins within the mucus layer.
Antibacterial activity is associated with mucus
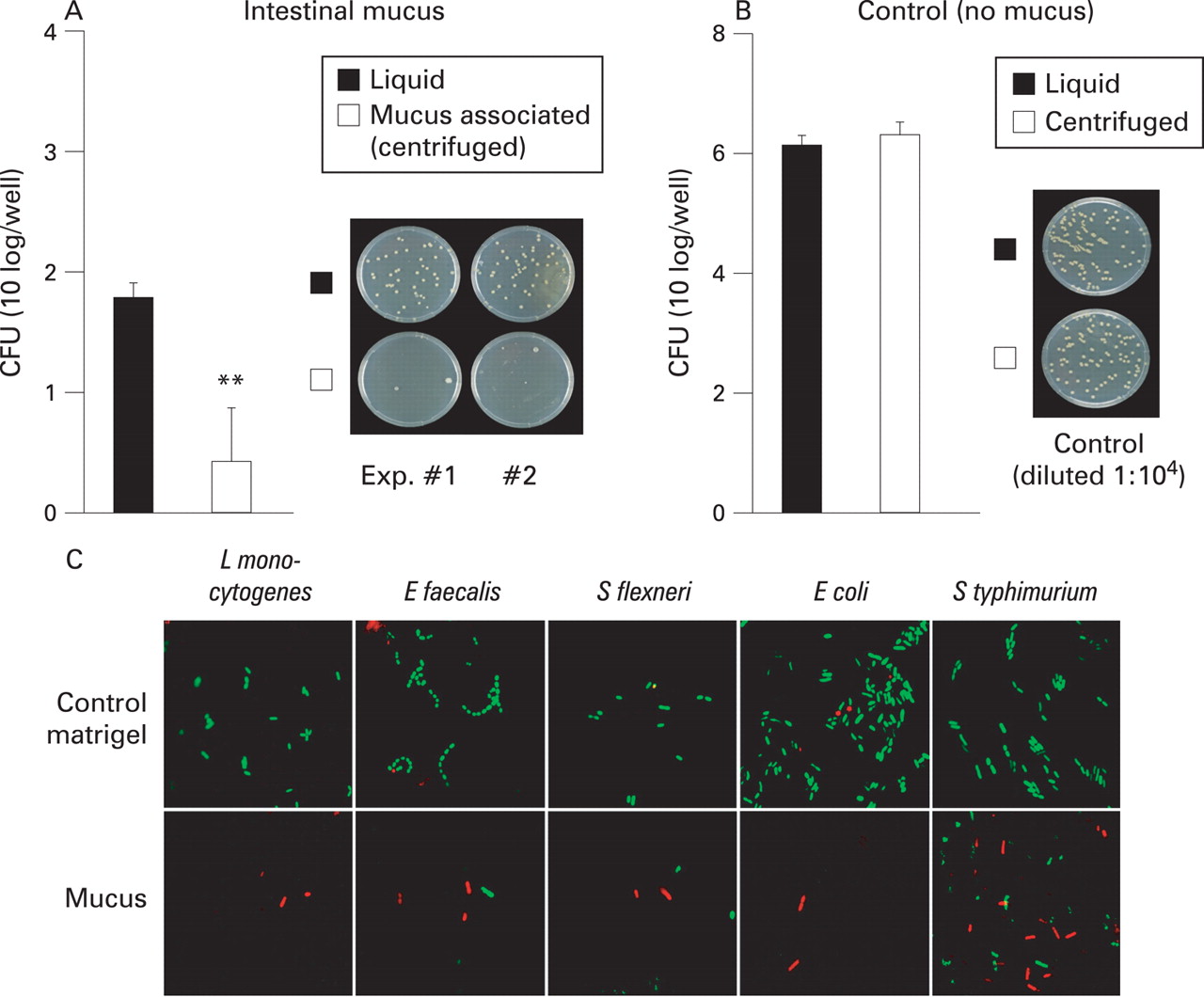
Isolated mucus from small intestine was used as a model to illustrate the protection provided by the overlaying mucus surface. Native isolated mucus contained relatively few viable aerobic bacteria (data not shown). Exogenous bacteria (106 colony-forming units (CFU)) were added on top of the mucus (fig 4A). Close contact between bacteria and the mucus barrier was modelled by forced sedimentation using centrifugation of the bacteria (mucus-associated bacteria) while the bacteria that were not subjected to centrifugation mimic microbes with a less close association with the mucus-covered epithelium (liquid bacteria). Interestingly, addition of bacteria to mucus-containing wells under both conditions (liquid bacteria and mucus-associated bacteria) killed off a significant portion of the bacteria, most probably due to components present or released from mucus (fig 4A,B). Importantly, killing efficiency was significantly (p<0.01) enhanced when the bacteria were brought into close contact with mucus (mucus associated). In fact, >99.999% of the added bacteria were killed when brought into close contact with mucus (fig 4A). A similar effect was observed starting with one log fewer bacteria, but was less evident when the bacterial load was increased by one log, suggesting a dose-dependent killing mechanism (data not shown). The increased killing was not induced by forced sedimentation, as CFU in the absence of mucus did not change after centrifugation (fig 4B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
To demonstrate clearly that the killing was mucus dependent, we performed experiments using an artificial gel “matrigel” as control and expanded the repertoire of test bacteria. The matrigel is a solubilised basement membrane preparation rich in laminin, collagen IV, heparan sulfate proteoglycans, and entactin. Live or dead staining indicated a dominant staining of live bacteria in contact with matrigel, as opposed to exclusively dead bacteria within the intestinal mucus layer (fig 4C). Also, fewer bacterial cells were observed within the mucus as compared with the matrigel, suggesting extensive bacterial lysis in the mucus. Interestingly, S Typhimurium, a bacterium that is known to have reduced susceptibility to antimicrobial peptide-mediated killing29 and to antimicrobial peptides (fig 1D,E) exhibited a higher proportion of viable cells as compared with the other bacteria tested (fig 4C).
The combined data show that the reduction in recovered bacteria (fig 4A) is not likely to be a consequence of trapped bacteria since the whole sample (including mucus) was used for plating and that mucus does not show the presence of notable trapped bacteria compared with artificial matrigel (fig 4C).
DISCUSSION
The intestine is colonised by a dense and highly dynamic microflora, which fulfils essential biological functions (30 and references therein). In addition, the enteric surface is constantly challenged by various enteropathogenic bacteria, which aim to penetrate the epithelial barrier to gain entry to the subepithelial space. Antimicrobial factors are probably major contributors to the mechanisms that confer a stable microflora, control bacterial overgrowth and prevent infection with enteroinvasive bacteria, thus facilitating the main organ function, namely the absorption of nutrients and fluid.
Defensins, CRS peptides, lysozyme P and phospholipase A2-IIA are released from Paneth cells of the small intestine so that sterilising conditions are achieved within the crypt compartment. Most of the antibacterial peptides and proteins that we identified here in whole small intestine or crypt preparations were also present in mucus preparations, but not in the intestinal lumen. The bactericidal components have been described to diffuse actively outwards driven by intestinal contractions, mixed with proteins from dying cells.31 We now show that the content is retained in close proximity to the epithelial layer by its association with the mucus layer. We further noted a clear contact-dependent killing of bacteria by intestinal mucus. This is in accordance with recent data suggesting that the initiation of biofilm formation occurs at the lumen–mucus interface and that the mucus is sparsely colonised.32 Our data explain this mechanism since the bacterial killing capacity of the antimicrobial peptides identified in the mucus is well known.33 The low antimicrobial activity of luminal extract probably results from poor diffusion through the mucus34 or possible degradation in the lumen. Defensins, however, are rather stable against trypsin degradation,35 speaking in favour of poor diffusion. These findings may reflect the close functional proximity of peptides and mucus in intestinal physiology and support the idea of the peptide-containing mucus layer as a combined physical and biochemical shield.
Mucus is produced by goblet cells and enterocytes, and a wide array of bioactive factors have been described to stimulate mucus production.36 The production of intestinal mucins parallels the postnatal bacterial colonisation (MWH, unpublished results) and the composition–thickness of the intestinal mucus was shown to be in part influenced by bacterial colonisation.36 The antibacterial peptide-enriched mucus shares certain functional similarities with the recently described neutrophil extracellular traps (NETs). NETs are a network of neutrophil DNA extruded from the dying neutrophil and complexed with enzymes and antimicrobial proteins.37 NETs similarly retain the biochemical antibacterial substances to provide high local concentrations of antimicrobial components that efficiently bind, disarm and kill microbes.
Mucus-associated antimicrobial components killed commensals as well as pathogens (figs 1 and 4). We speculate that the antimicrobial activity limits growth of mucus-associated commensals as well reduces the establishment of pathogens. A certain commensal community, however, is necessary for proper development of the mouse small intestine.38–40 This is in agreement with recent data from us and others41 42 that show a developmental switch of antibacterial peptide production around the time of weaning and that fewer α-defensin peptides are produced early in life. This would allow closer contact of commensals and the epithelial layer before weaning and have an impact on intestinal development.
Interestingly, the mucus barrier in patients suffering from Crohn’s disease seems to be distorted, leading to direct contact of commensal bacteria with the epithelial lining, which was not the case in healthy individuals.14 43 Furthermore, patients with Crohn’s ileitis have reduced α-defensin levels.12 13 The opposite situation is exemplified by transgenic mice expressing an extra human defensin, which were more resistant against enteric Salmonella infection and in addition displayed an altered composition of the intestinal microflora.11–13 Taken together, these findings show that α-defensins plays a central role in the enteric antibacterial defence, and disturbance of the function of antimicrobial peptides or mucus production may significantly impair the host intestinal barrier and lead to enhanced susceptibility to enteric infections or inflammatory conditions.
We did not find any prominent β-defensin expression in the mouse small intestine although β-defensins have been shown to play an important role in human colon.44 Most β-defensins analysed have low expression in non-inflamed tissue, except mBD-1, and are not prominently expressed in normal mouse colon.45 46 So far β-defensins have not been isolated from colon at the protein level. This as well as other factors contributes to a higher colonisation in the colon than in the small intestine, which produces high amounts of α-defensins.
Strikingly, transgenic mice with a temporal depletion of functional Paneth cells are healthy and show no major signs of enteric microbial overgrowth.47 One potential source of protection is epithelial cells that are constantly renewed and shed from the villus tip. Here, we identified several ribosomal proteins (ubiquicidin, L26, L28, L29 and L35) with significant antibacterial activity in mucus preparations. These proteins may act synergistically with classic antimicrobial peptides/proteins (eg, defensins, lysozyme P and phospholipase A2-IIA) and broaden the antibacterial spectrum. Most of these components have been detected in conventionally reared and germ-free mice.48 Although the measured activity cannot conclusively be ascribed to the ribosomal proteins, other reports have previously identified ribosomal proteins with antimicrobial activity.49–51 Ribosomal proteins are classically found as intracellular, but there is, however, increasing evidence that some individual ribosomal proteins may have extra-ribosomal functions.52 Together, these findings suggest that the high cell turnover in the small intestine53 might contribute to antibacterial defence by the release of cytosolic proteins such as ribosomal proteins.
In conclusion, our results demonstrate that the composition of antimicrobial compounds in the mucus resembles the spectrum found in isolated crypt tissue. Secreted antimicrobial peptides may therefore not diffuse into the gut lumen but rather be retained in the mucus layer to form a physical and biochemical barrier. This would ensure high local peptide concentrations and efficient and rapid killing of approaching microbes in the absence of significant interference with the luminal physiological microflora.
Acknowledgments
We thank E Cederlund for performing the Edman degradation, and Dominique Gütle for excellent technical work. This work was supported by grants of the Swedish Research Council (KP, MWH, MA and BHN), SSAC (MA), The Swedish Foundation for International Cooperation in Research and Higher Education (MA), Ruth och Richard Juhlins stiftelse (KP), Cancerfonden (MWH), Swedish Royal Academy of Sciences (BH-N), Torsten and Ragnar Söderbergs foundation (BH-N), Swedish Foundation for Strategic Research (BH-N) the Deutsche Forschungsgemeinschaft Me2037/2-1 (UM-H) and Ho 2236/5-2 (MH).
REFERENCES
Footnotes
-
Competing interests: None.
Linked Articles
- Digest